



2. Genética mendeliana.
2. Genética mendeliana.
La genética mendeliana es la parte de la genética que sigue la metodología que ideó Mendel. Se basa en el estudio de las proporciones en las que se heredan las características de los individuos.
Se considera a Mendel como fundador de la genética, aunque la comunidad cien- tífica no tuvo en cuenta su obra hasta 40 años más tarde, cuando sus trabajos fueron redescubiertos independientemente por De Vries, Correns y Von Tschermak.
Durante las dos terceras partes del siglo xx, se ha podido descubrir la función de muchos genes, las leyes que rigen su transmisión hereditaria, se ha evaluado matemáticamente la probabilidad de heredar una determinada característica, se ha mejorado el rendimiento de muchos cultivos, en épocas en las que la naturaleza íntima de los genes no era aún accesible al investigador.
A la luz de los conocimientos actuales, podemos analizar las posibilidades que nos brinda el estudio de las características hereditarias de la descendencia de un cruza- miento.
El éxito de los trabajos de Mendel se debe a varios factores:
- La selección adecuada del material de partida: la planta del guisante.
- El riguroso estudio estadístico de la descendencia, aspecto que no tuvieron en cuenta los biólogos anteriores.
- La simplificación del problema, al analizar un solo carácter de los muchos que se podían encontrar alterados.
2.1. Conceptos básicos de la herencia biológica.
Existen unos conceptos fundamentales en Genética que permiten la adecuada comprensión de los mecanismos hereditarios. Son los siguientes:
- Genética. Ciencia que estudia la transmisión de los caracteres hereditarios.
- Carácter hereditario. Característica morfológica, estructural o fisiológica presente en un ser vivo y transmisible a la descendencia.
- Gen. Término creado por Johannsen en 1909 para definir la unidad estructural y funcional de transmisión genética. En la actualidad, se sabe que un gen es un fragmento de ADN que lleva codificada la información para la síntesis de una determinada proteína. Mendel denominó “factor hereditario”.
- Genotipo. Conjunto de genes que posee un individuo.
- Fenotipo. Características que muestra un individuo, es decir, expresión externa del genotipo.
- Alelos. Término introducido por Bateson en 1902 para indicar las distintas formas que puede presentar un determinado gen.
- Homocigoto o raza pura. Individuo que posee dos alelos idénticos para el mismo carácter.
- Heterocigoto o híbrido. Individuo que tiene dos alelos distintos para el mismo carácter.
- Gen o alelo dominante. Gen cuya presencia impide que se manifieste la acción de otro alelo distinto para el mismo carácter.
- Gen o alelo recesivo. Gen que sólo manifiesta su acción en ausencia de un alelo dominante, es decir, únicamente aparece en el fenotipo si se encuentra en homocigosis. - Genes o alelos codominantes. Alelos para el mismo carácter que poseen idéntica capacidad para expresarse y, cuando se encuentran juntos en el mismo individuo, éste manifiesta la acción de ambos.
- Cromosomas homólogos. Pareja de cromosomas en células diploides, que procede uno del progenitor paterno y el otro del materno, son iguales morfológicamente (excepto los cromosomas sexuales) pero no son idénticos, puesto que no tienen la misma composición química, al contener diferentes genes alelos uno y otro cromosoma.
- Locus. Lugar ocupado por un gen en un cromosoma. El plural es loci por ser palabra latina.
- Herencia dominante. Es aquella en la que hay un alelo, el llamado dominante, que no deja manifestarse al otro, el llamado alelo recesivo
- Herencia intermedia. Es aquella en la que uno de los alelos muestra una dominancia incompleta sobre el otro. Así pues, los híbridos tienen un «fenotipo intermedio» entre las dos razas puras.
- Herencia codominante. Es aquella en la que los dos alelos son equipotentes, y por tanto no hay dominancia. Los híbridos presentan las características de las dos razas puras a la vez.
- Dihíbridos. Son los individuos con heterocigosis en dos pares de genes.
- Polihibridos. Son los seres con heterocigosis para muchos pares de genes.
- Alelos letales. Son aquellos alelos que poseen una información deficiente para un carácter tan importante que, sin él, el ser muere. Los alelos letales pueden producir la muerte a nivel del gameto o a nivel del cigoto, pudiendo suceder entonces que el individuo no llegue a nacer, o bien que muera antes de alcanzar la capacidad reproductora. Los alelos letales suelen ser recesivos, por lo que necesitan darse en homocigosis para manifestarse.
- Cariotipo. Conjunto de cromosomas de un individuo, característico de cada especie en cuanto a forma, tamaño y número, que se perpetúan en la descendencia.
- Simbología. Los genes se simbolizan con letras. Si es herencia dominante y sólo hay dos alelos, el dominante se representa con mayúscula y el recesivo con minúscula. La letra escogida puede ser la inicial del nombre del carácter dominante o la del carácter recesivo.
Otro tipo de notación, que permite además simbolizar más de dos alelos, es el uso de exponentes (superíndices). Un caso en el que se utiliza esta anotación es en la herencia de los grupos sanguíneos humanos ABO.
2.1.1. Genotipo y fenotipo.
Los caracteres que manifiesta un individuo es indudable que reconocen como causa inicial el factor hereditario, pero a medida que dicho individuo se desarrolla, también los factores del medio ambiente (clima, alimentación, higiene, etc.) dejan sentir su influencia. En consecuencia, todo carácter depende de dos tipos de factores:
a) Heredables o genéricos, recibidos de los progenitores a través de las células reproductoras de estos y, por tanto, internos.
b) No heredables, procedentes del medio ambiente y, por tanto, externos, que pueden influir a lo largo de la vida del individuo imprimiendo modificaciones a los caracteres heredados.
En conclusión, todo carácter depende de la acción combinada y recíproca entre los factores hereditarios y los ambientales.
Como consecuencia es lógico pensar, que los caracteres no siempre son un fiel reflejo de los factores hereditarios, es decir, que la imagen que observamos en un individuo es el resultado de los factores heredados y de la acción que recibe del medio ambiente.
Para poder establecer esta distinción se han introducido los términos genotipo y fenotipo .
El genotipo es el conjunto de factores hereditarios que posee un individuo por haberlos recibido de sus progenitores. El fenotipo es el aspecto observable cuyo aspecto ha sido adquirido como consecuencia del genotipo que posee y de la acción del medio ambiente, o dicho de otro modo, el fenotipo es la manera de manifestarse el genotipo después de haber actuado sobre él los factores ambientales. Además, como veremos más adelante, existen factores heredables que no llegan a manifestarse (los llamados factores recesivos) y, por tanto, no pueden ser apreciados, lo que también influye en que el fenotipo no refleje fielmente al genotipo.
Mientras el genotipo es estable durante toda la vida, el fenotipo va cambiando. Así, cualquier ser vivo posee siempre los mismos factores hereditarios, pero su aspecto exterior varía profundamente de la edad joven a la adulta o en la vejez.
De todo lo dicho resulta, que lo único heredable es el genotipo, ya que la influencia que ejercen los factores ambientales sobre los caracteres, sólo afectan al individuo sobre el que actúan, perdiéndose con él sin transmitirse a sucesivas generaciones.
2.2. Las leyes de Mendel.
Los descubrimientos de Mendel pueden resumirse en tres leyes, que constituyen los fundamentos básicos de transmisión genética. La terminología que empleo Mendel es de difícil comprensión; por ello vamos a utilizar la terminología actual.
2.2.1. Primera ley de Mendel.
Llamada también ley de la uniformidad de los híbridos de la primera generación, dice que: cuando se realiza el cruzamiento entre dos individuos de la misma especie pertenecientes a dos variedades o razas puras (homocigóticos) todos los híbridos de la primera generación filial son iguales.
En la actualidad esta ley expresa así. “El cruce de dos razas puras da un descendencia híbrida uniforme tanto fenotipica como genotipicamente.”
Esta uniformidad de todos los individuos de la F1 puede manifestarse, bien por parecerse a uno de los padres (herencia dominante), bien porque aparezca un fenotipo con aspecto intermedio (herencia intermedia). Veamos seguidamente un ejemplo de cada caso.
Empecemos por la herencia dominante
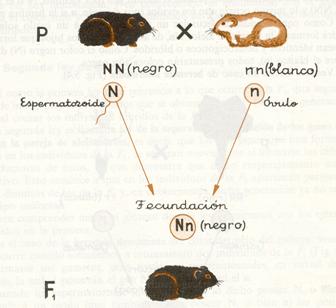
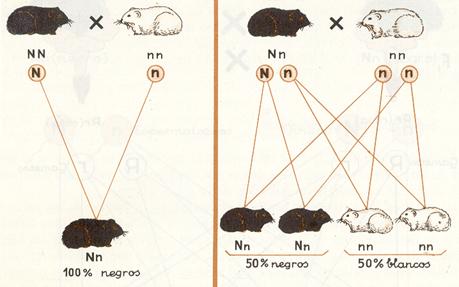
Esquema de la primera ley de Mendel en dominancia.
Si cruzamos un cobayo (conejillo de Indias) homocigótico para el color negro del pelo (NN ) con otro también homocigótico para el color blanco (nn), todos los cobayos que se obtengan de este cruzamiento serán de color negro (Nn), ya que este domina sobre el blanco.
La explicación de este resultado queda claramente expresada en el esquema. Cuando los individuos homocigóticos que se cruzan (generación P ) forman sus células reproductoras (espermatozoides en el macho y óvulos en la hembra), en virtud del fenómeno de la meiosis los genes que forman la pareja de alelomorfos y que se hallan situados en los respectivos cromosomas homólogos, se separan, yendo a parar cada uno de ellos a una célula reproductora. Como los dos genes que forman la pareja son iguales (NN o bien nn) es lógico que todos los gametos posean el mismo gen (por ejemplo, N si el macho era NN) y lo mismo ocurre con los óvulos (por ejemplo, n si la hembra era nn).
Como consecuencia, al fecundar un espermatozoide a un óvulo solamente podrá formarse la pareja de alelos Nn , de ahí que todos los hijos que forman la F1sean idénticos y heterocigóticos o híbridos. Como el color negro (N) domina sobre el blanco (n), todos presentarán coloración negra.
Veamos ahora un caso de herencia intermedia.
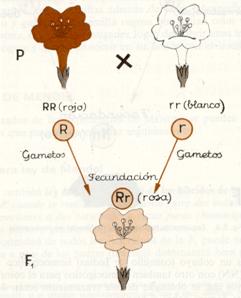
Esquema de la primera ley de Mendel en herencia intermedia.
Existen dos variedades de la planta «dondiego de noche» (Mirábilis jalapa) que se diferencian por el color de sus flores: en unas, rojo; en otras, blanco. Si cruzamos una planta homocigótica para el color rojo (RR ), con otra también homocigótica para el color blanco (rr) todas las plantas que se obtengan de este cruzamiento serán de color rosa (Rr).
Como puede observarse en el esquema, la interpretación de los resultados es la misma que en el caso anterior, con la única diferencia que el fenotipo de las flores de la F1 no corresponde a ninguno de los de las plantas progenitoras porque no hay dominancia, y en consecuencia se manifiestan con la misma eficacia el color rojo y el blanco, resultando de ello un color rosa intermedio entre ambos.
2.2.2. Segunda ley de Mendel.
Así como la primera ley hace referencia a lo que ocurre en la Fl, esta segunda trata de interpretar los resultados que se obtienen en la F2 (segunda generación filial) al cruzar los individuos híbridos de la Fl.
La segunda ley es llamada ley de la separación o disyunción de los genes que forman la pareja de alelomorfos, es decir, que los dos genes que han formado pareja en los individuos de la Fl, se separan nuevamente al formarse las células reproductoras de éstos, lo que demuestra que dicho emparejamiento no es definitivo. Esto conduce a que en los individuos de la F2 aparezcan parejas de alelos distintos de los de la Fly, en consecuencia, dicha generación ya no es de genotipo uniforme.
Así, puede formularse esta ley actualmente: “Al cruzar entre sí los híbridos obtenidos en la primera generación, los caracteres antagónicos que poseen se separan y se reparten entre los distintos gametos, apareciendo así varios fenotipos en la descendencia”
Para comprender mejor el alcance de esta ley, seguiremos con los ejemplos expuestos en la primera.
En el caso de la herencia dominante del color del pelo del cobaya, veamos qué ocurre cuando sometemos a cruzamiento dos individuos de la Fl. Al formarse sus gametos, sean óvulos o espermatozoides, en virtud de la meiosis, la mitad poseerán el gen N y la otra mitad el n.
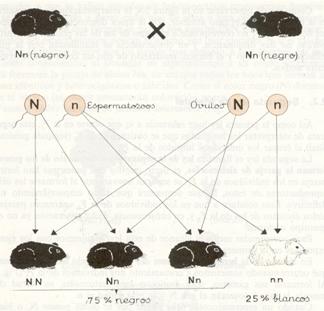
Representación esquemática de la segunda ley de Mendel en dominancia.
Cuando un espermatozoide (que puede por lo dicho poseer N , o bien n) fecunde a un óvulo (que también puede poseer N, o bien n) las posibles combinaciones para formar la pareja de alelos (genotipo) en los individuos de la F2 serán NN, Nn, Nn y nn,según se expresa en el esquema. Así pues, en la segunda generación filial (F2) se presentará una proporción genotípica de 1 : 2 : 1, es decir, que de 100 individuos, el 25 por 100 serán de genotipo NN; el 50 por 100, de genotipo Nn, y el 25 por 100, de genotipo nn. Pero como se trata de un caso de herencia dominante, el fenotipo de los individuos NN y Nn será igual, de ahí que la proporción fenotípica será 3: 1, es decir, el 75 por 100 de cobayos negros y el 25 por 100 de blancos.
Como vemos, los genes N y n que estaban reunidos en los individuos de la Fl se separan para dar nuevas combinaciones, concretamente NN y nn iguales a las de sus abuelos (generación P) que habían desaparecido en la Fl, lo que indica, como dice la ley, que los alelomorfos de los individuos de la Flpueden separarse.
Si ahora examinamos lo que ocurre en los casos de herencia intermedia , tal como la que presenta el dondiego de noche, los resultados serán los siguientes:
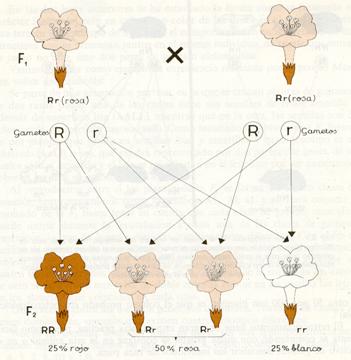
Representación esquemática de la segunda ley de Mendel en herencia intermedia.
En la F2 los genotipos se hallarán también en la proporción 1 RR : 2 Rr: 1 rr, pero como aquí no hay herencia dominante, la proporción de fenotipos será la misma que la de genotipos, es decir, un 25 por 100 de flores rojas, un 50 por 100 de flores rosas y un 25 por 100 de flores blancas.
2.2.3. Cruzamiento prueba y retrocruzamiento.
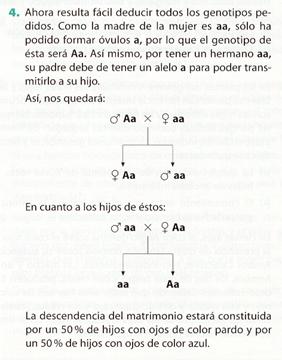
Recibe el nombre de retrocruzamiento el cruzamiento entre un individuo y uno de sus parentales. Cuando el parental utilizado es el homocigótico recesivo, se denomina cruzamiento prueba, ya que con este método se puede averiguar si un individuo es homocigótico dominante o heterocigótico. (Muchos autores no distinguen entre retrocruzamiento y cruzamiento prueba)
Si todos los descendientes del cruzamiento prueba son del fenotipo dominante; el individuo problema debe ser, necesariamente, homocigótico.
Por el contrario, si la mitad de la descendencia presenta el fenotipo dominante y la otra mitad es recesivo, el individuo problema es heterocigótico.
Si al cruzar un cobayo de fenotipo negro (individuo problema) con otro de fenotipo blanco (cruzamiento prueba), toda la descendencia es de color negro, es que el cobayo probado era de genotipo puro para el color negro del pelo (NN ); pero si la mitad de la descendencia manifiesta pelaje negro y la otra mitad blanco, es que el cobayo probado (individuo problema) era heterocigótico o híbrido para el pelo negro (Nn).
2.2.4. Tercera ley de Mendel.
Llamada ley de la herencia independiente de los caracteres, porque expresa el hecho de que cada uno de los caracteres hereditarios se transmite a la descendencia con absoluta independencia de los demás.
Hoy se enuncia esta ley así: “Los distintos caracteres no antagónicos se heredan independientemente unos de otros, combinándose al azar en la descendencia”
En las dos leyes anteriores se ha estudiado la forma como se transmite un carácter (color del pelo en cobayas o color de las flores en el dondiego); pero esta tercera ley se ocupa de averiguar el comportamiento en la herencia de dos caracteres que se presentan juntos en el mismo individuo, de suerte que entran en juego no uno, sino dos pares de genes o alelomorfos (dihibridismo ).
Vamos a tomar como ejemplo la experiencia realizada por el propio Mendel sobre los guisantes:

Esquema de la tercera ley de Mendel.
Se parte de una generación paterna, en la que se cruzan plantas de guisantes de dos razas puras, una de las cuales tiene sus semillas de color amarillo y además de superficie lisa (AALL ), mientras que en la otra, las semillas son de color verde y superficie rugosa (aall). Como resultado se obtiene una Fl formada por plantas que producen semillas lisas y amarillas y cuyo genotipo es un dihíbrido (AaLl), o sea, que vendrá representado por dos parejas de genes, de los cuales sólo el amarillo y el liso se manifiestan en el fenotipo por ser dominantes sobre el verde y el rugoso.
Al reproducirse entre sí las plantas de la Flse formarán cuatro clases de gametos, tanto masculinos como femeninos: AL, Al, aL y al. Para conocer el resultado de la F2 basta tener en cuenta que cada clase de gameto masculino puede unirse durante la fecundación a cada una de las clases de gametos femeninos, lo que da lugar a dieciséis combinaciones diferentes. Con el fin de facilitar la escritura de estos dieciséis genotipos, se puede construir el llamado tablero de Punnett, colocando en la línea horizontal superior los cuatro tipos de gametos de un sexo y en la columna de la izquierda los cuatro de otro sexo, y como una tabla de doble entrada anotar en las casillas las letras de los gametos que coinciden en cada caso.
Observemos ahora los dieciséis genotipos obtenidos. Teniendo en cuenta los genes dominantes, veremos que existen cuatro fenotipos diferentes:
Nueve de semillas amarillas lisas , es decir, los dos fenotipos dominantes (cuando haya por lo menos un gen A y uno L, como ocurre en las casillas 1, 2, 3, 4, 5, 7, 9, 10 y 13).
Tres de semillas amarillas y rugosas , es decir, con el dominante del color y el recesivo del otro carácter (cuando haya uno o dos genes A, pero ninguno L, como ocurre en las casillas 6, 8 y 14).
Tres de semillas verdes y lisas , es decir, con el otro dominante (cuando haya uno o dos genes L, pero ninguno A, como ocurre en las casillas 11, 12 y 15).
Una de semillas verdes y rugosas , porque no hay ningún gen dominante para ninguno de los dos caracteres. Es el caso del genotipo de la casilla 16.
Como consecuencia, la proporción numérica entre los cuatro fenotipos será 9:3:3:1.
Si en el esquema consideramos las casillas diagonales 1, 6, 11 y 16, veremos que corresponden a los individuos homocigóticos o puros. De ellos el 1 y el 16 repiten el genotipo de los abuelos (AALL y aall), pero la 6 y la 11 representan nuevas combinaciones homocigóticas. De todo ello se deduce que en la herencia los caracteres liso-amarillo no permanecen siempre unidos, y lo mismo ocurre con los caracteres rugoso-verde, lo cual demuestra que «existe una independencia» entre ellos, ya que pueden formar combinaciones liso-verde y rugoso-amarillo.
Las excepciones a la tercera ley
La transmisión independiente de los caracteres no siempre se cumple, es decir, que muchos de ellos se transmiten juntos en la herencia. La explicación a esta excepción de la tercera ley de Mendel, se comprende fácilmente considerando que al estar localizados los genes en los cromosomas puede ocurrir que dos alelomorfos que rigen sendos caracteres se hallen situados en la misma pareja de cromosomas homólogos. Esto es muy fácil que ocurra si se tiene en cuenta que las parejas de genes son bastante más numerosas que los pares de cromosomas homólogos, por lo que cada una de estas parejas forzosamente debe contener un gran número de alelos.
2.3. Ejemplos de herencia mendeliana en animales y plantas.
La leyes de Mendel se cumplen siempre (salvo lo dicho para la tercera ley), pero el concepto de herencia mendeliana se suele reservar para aquellos casos sencillos, como los descritos hasta aquí, en que un carácter viene regido por un solo gen con dos variedades alélicas.
Podemos citar algunos ejemplos de herencia mendeliana en los animales y en el hombre.

En las plantas podemos citar el color de las flores, longitud del tallo, forma de las hojas, entre otros muchos ejemplos.
|
|
|

2.4. Teoría cromosómica de la herencia.
En la época que siguió a los experimentos de Mendel, segunda mitad del siglo XIX, no se conocía donde se ubicaban, dentro de los seres vivos, los factores hereditarios de los que hablaba Mendel y que eran responsables de los resultados experimentales que obtuvo.
A partir de 1870 se avanzó en el estudio de la célula, de modo que se observaron los cromosomas en núcleos celulares y que durante la división celular los cromosomas se dividían longitudinalmente en dos, los cuales pasaban a los núcleos de las células hijas.
También se comprobó en esta época histórica que el número de cromosomas es constante para cada especie y que cuando se forman las células reproductoras o gametos, dicho número queda reducido a la mitad (reducción meiótica), restableciéndose su número al unirse los dos gametos en la fecundación.
No pasó mucho tiempo hasta percibir la correlación que existe entre el comportamiento de los cromosomas y el de los factores hereditarios de Mendel, aunque el número de estos sea grande y el de aquellos limitado para cada especie.
En 1902, Sutton, propuso la siguiente hipótesis: los factores hereditarios debían de estar localizados en los cromosomas.
Al norteamericano y premio Nóbel Thomas H. Morgan le correspondió contrastar la validez de la hipótesis de Sutton, con la cual, por cierto, no se mostraba de acuerdo. Consiguió verificarla y demostró además que los genes se hallan ordenados linealmente en los cromosomas. Morgan utilizó para ello una pequeña mosca que tiene un ciclo de desarrollo corto y es fácil de criar en grandes cantidades y en poco espacio, es la mosca de la fruta, mosca del vinagre o drosófila (Drosophila melanogaster). Presenta pocos cromosomas, sólo 10, cuatro parejas y muy distintos entre sí.
Los resultados de los experimentos llevaron a Morgan a formular una serie de conclusiones que conforman la teoría cromosómica de la herencia:
-Los factores hereditarios (o genes) que determinan las características fenotipicas de los organismos se localizan en los cromosomas.
-Cada factor (o gen) ocupa un lugar específico en un cromosoma determinado. A este lugar se le denomina locus (en plural loci). En los organismos diploides cada factor está duplicado (hoy se llaman alelos) y se sitúan en un mismo locus de un par de cromosomas homólogos.
-Los factores (o genes) o loci se disponen linealmente a lo largo de los cromosomas.
2.4.1. Los genes y los cromosomas.
Gracias a los trabajos realizados por T. H. Morgan con la mosca de la fruta (Drosophila melanogaster) se observó que la tercera ley o "principio de independencia de los caracteres" no se cumplía siempre. Se cumple cuando las parejas alélicas que rigen dos caracteres hereditarios se encuentran situadas en diferentes cromosomas.
Cada especie animal o vegetal tiene muchos más pares de genes que pares de cromosomas. Obviamente, debe haber muchos genes en cada cromosoma. Así, por ejemplo, el hombre tiene 23 pares de cromosomas, algunos grandes y otros pequeños, pero millares de genes. Los cromosomas se heredan como unidades y, por tanto, los genes de cualquier cromosoma tienden a ser heredados juntos. Aquellos genes localizados en el mismo cromosoma y que, por consiguiente, se transmiten juntos a la descendencia se denominan genes ligados .
La existencia de genes ligados fue demostrada por Morgan en el año 1911, investigando la herencia del color del cuerpo y de cierta anomalía de las alas, denominadas alas vestigiales o reducidas, de la mosca de la fruta Drosophila melanogaster. Realizó el siguiente cruzamiento:
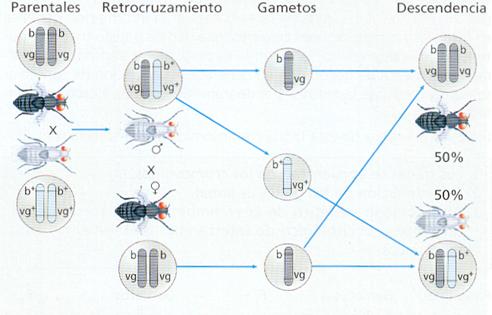
Todos los individuos de la F1, según la primera ley de Mendel, eran iguales, de genotipo heterocigoto (b+b vg+vg ) y de cuerpo gris y alas normales, lo cual significa que estos caracteres (color del cuerpo gris y alas normales) son dominantes.
En la descendencia del retrocruzamiento de un macho de esta F1 con una hembra doble recesiva (bb vgvg ) no aparecía ningún individuo de cuerpo gris y alas vestigiales, ni de cuerpo negro y alas normales, tal como hubiese ocurrido si la tercera ley de Mendel se cumpliese. Estos resultados llevaron a Morgan a deducir que los genes determinantes del cuerpo color negro y de las alas vestigiales se transmiten juntos en el mismo cromosoma y, por tanto, son genes ligados.
2.4.2. La meiosis y su relación con las leyes de Mendel.
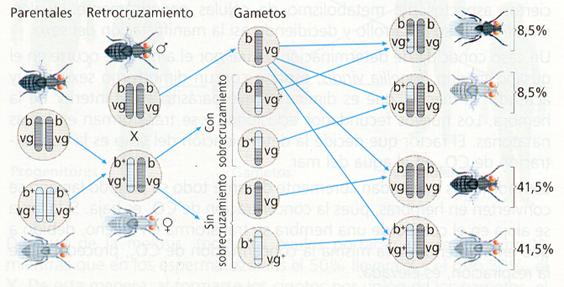
Los resultados obtenidos en el experimento de Morgan eran diferentes si en el retrocruzamiento se utiliza una hembra de genotipo doble heterocigoto (b+b vg+vg ) a la que se cruza con un macho de genotipo doble homocigoto recesivo (bb vgvg). La descendencia obtenida estaba formada por:
- 41,5 % de cuerpo gris y alas normales.
- 41,5 % de cuerpo negro y alas vestigiales.
- 8,5 % de cuerpo negro y alas normales.
- 8,5 % de cuerpo gris y alas vestigiales.
Si los genes que determinan el color del cuerpo y la forma de las alas están ligados no es posible que se obtengan estos resultados, y si no lo estuviesen, según las leyes de Mendel, se habría obtenido un 25% de cada uno de los cuatro fenotipos posibles. Estos genes no son completamente independientes, pero tampoco se encuentran completamente ligados. La explicación a esta aparente contradicción fue dada por el propio Morgan, al suponer que los cromosomas en los que se localizan los genes se intercambian fragmentos, proceso que se denomina recombinación genética .
Durante la meiosis, en la profase I, cuando las cromátidas homólogas se encuentran estrechamente unidas, se entrecruzan e intercambian, material genético (sobrecruzamiento ). El punto concreto donde se entrecruzan se llama quiasma y al intercambio de material genético se denomina recombinación genética.
Estas experiencias confirmaron plenamente la teoría cromosómica de la herencia, haciendo compatible la tercera ley de Mendel con la agrupación de miles de genes ligados en un solo cromosoma.
Los mapas cromosómicos muestran el orden lineal de los genes en los cromosomas. En la especie humana se está llevando a cabo gracias al denominado Proyecto Genoma Humano, uno de cuyos objetivos es determinar la ubicación exacta de todos y cada uno de los miles de genes que posee el hombre.
2.5. Determinismo del sexo y herencia ligada al sexo.
DETERMINACIÓN DEL SEXO
Un aspecto importante del fenotipo de un organismo es su sexo, que en la mayoría de los casos se halla controlado por genes, situados en cromosomas específicos (sexuales), aunque también existen organismos en los que depende de factores ambientales.
Determinación cromosómica
En numerosos organismos, entre ellos la especie humana, la determinación del sexo se realiza por la presencia de cromosomas especiales llamados sexuales o heterocromosomas, que se diferencian del resto, que son los autosomas o cromosomas autosómicos.
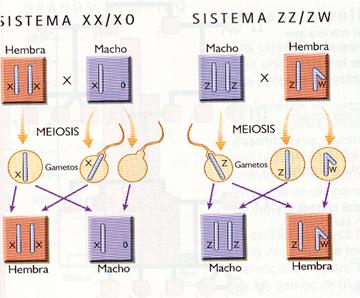
· Sistema XX/XY. Es el tipo de determinación de la especie humana y de otros muchos animales (mamíferos, equinodermos, moluscos y gran número de artrópodos). Los cromosomas sexuales se denominan X e Y, en función de su forma. Las hembras tienen una dotación XXy son de sexo homogamético, ya que todos los gametos que producen llevan el cromosoma X, mientras que los machos son XY, es decir, de sexo heterogamético, puesto que la mitad de los gametos producidos portan el cromosoma X y la otra mitad el Y.
· Sistema XX/XO o ZZ/ZO. Determinación propia de algunos insecto caracterizados por que uno de los dos sexos solo tiene un cromosoma sexual.
· Sistema ZZ/ZW.Tipo de determinación propia de las aves, de algunos anfibios y reptiles y de los lepidópteros (mariposas). Se utiliza la notación ZZ/ZW para no confundirse con la determinación XX/XY; ya que en este sistema las hembras son el sexo heterogamético (ZW), mientras que los machos son el sexo homogamético (ZZ).
Determinación por haplodiploidía
En algunos grupos de insectos sociales (abejas, avispas y hormigas) no existen cromosomas sexuales. El sexo se halla determinado por el número de dotaciones cromosómicas; así, los individuos diploides (2n) son hembras y los haploides (n) son machos. Las hembras se desarrollan a partir de óvulos fecundados, mientras que los machos lo hacen a partir de óvulos sin fecundar.
En el caso de las abejas, la reina, con dotación cromosómica diploide, posee óvulos, que si no son fecundados desarrollarán por partenogénesis machos o zánganos, mientras que si son fecundados se originarán hembras, una de las cuales será alimentada con jalea real y se convertirá en la reina, única hembra fértil de la colmena.

Determinación génica
En algunas plantas, y también en algunos animales, el que un individuo sea de un sexo o sea de otro depende de una o de varias parejas de genes. Muy estudiado es el caso de una planta, la Ecballíum elateríum (pepinillo del diablo), en la que la determinación del sexo depende de una serie alélica (aD > a+ > ad), en donde el alelo aD determina la masculinidad (plantas con genotipos aDaD, aDa+ y aDad), el alelo a+ determina características de hermafroditismo (plantas con genotipos a+a+, a+ad) y el alelo ad determina características femeninas (genotipo adad).
Determinación ambiental
En algunos animales, la determinación del sexo depende de circunstancias ambientales. Factores externos influyen en ciertos aspectos del metabolismo de células genéticamente idénticas, modificando su desarrollo y decidiendo así la manifestación del sexo. Entre los grandes saurios (cocodrilos, aligatores y caimanes) el sexo está determinado por la temperatura a la que se incuban los huevos: si es superior a los 27 ºC se obtendrán machos y si es inferior nacerán hembras.
HERENCIA LIGADA AL SEXO
Thomas Hunt Morgan fue uno de los primeros científicos en relacionar la herencia de algunos caracteres con el sexo, al descubrir que el gen causante del color de ojos en la mosca Drosophila melanogaster se encuentra localizado en el cromosoma X. De esta forma se demuestra que estos cromosomas no solo llevan los genes que determinan el sexo, sino otros que influyen sobre caracteres hereditarios no relacionados con el mismo.
A partir de este descubrimiento se habla estrictamente de caracteres ligados al sexo como aquellos que están determinados por genes localizados en los cromosomas sexuales; se trata de caracteres que aparecen en uno solo de los sexos o bien, si lo hacen en ambos, con más frecuencia en uno de ellos que en el otro.
Herencia ligada al sexo en la especie humana
La especie humana tiene 46 cromosomas, es decir, 22 parejas de autosomas más una pareja de cromosomas sexuales, XX en la mujer y XY en el hombre. Como en otras muchas especies, el cromosoma X es mayor que el del Y, pero en ambos existe un largo segmento homólogo, que les permite aparearse y entrecruzarse durante la meiosis, y un corto segmento diferencial, no apareable, con genes específicos para cada uno de los dos cromosomas.
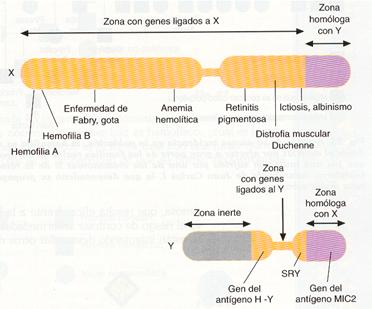
Esquema de los cromosomas X e Y humanos, indicando la localización de algunos genes ligados al sexo.
- Herencia ligada al cromosoma Y. Todos los genes que se encuentran en el segmento diferencial del cromosoma y son heredados únicamente por los hijos varones. Se llama herencia holándrica, ya que se manifiesta solo en los varones, como la presencia de pelos en las orejas o la ictiosis, enfermedad de la piel caracterizada por la formación de escamas y cerdas.
- Herencia ligada al cromosoma X. Dado que el número de genes, y por tanto el de caracteres ligados al segmento diferencial del cromosoma X, es más numeroso que el de los ligados al Y; se generaliza como herencia ligada al sexo toda la que se encuentra ligada al X. Tal es el caso del daltonismo y de la hemofilia, enfermedades provocadas por un gen recesivo situado en el segmento diferencial del cromosoma X. Debido a esta ubicación, para que una mujer padezca la enfermedad debe ser homocigoto recesivo, mientras que en los hombres, que son hemicigotos, basta para que la padezcan que el gen se encuentre en el único cromosoma X que tienen.
El daltonismo es un defecto visual que hace que la persona afectada tenga dificultades para distinguir con claridad el color rojo del color verde.

La hemofilia es una enfermedad que provoca problemas en la coagulación de la sangre debido a la carencia de algunos de los factores proteicos responsables de la misma.
Herencia influida por el sexo
Existen caracteres, como la calvicie en la especie humana y la presencia o ausencia de cuernos algunas razas ovinas, que están determinados por genes situados en la parte homóloga de los cromosomas sexuales o bien en autosomas, y cuya manifestación depende del sexo.
