



Metabolismo 4 (soluciones)
1.- Interprete el esquema adjunto. ¿Cuál es su significado metabólico?
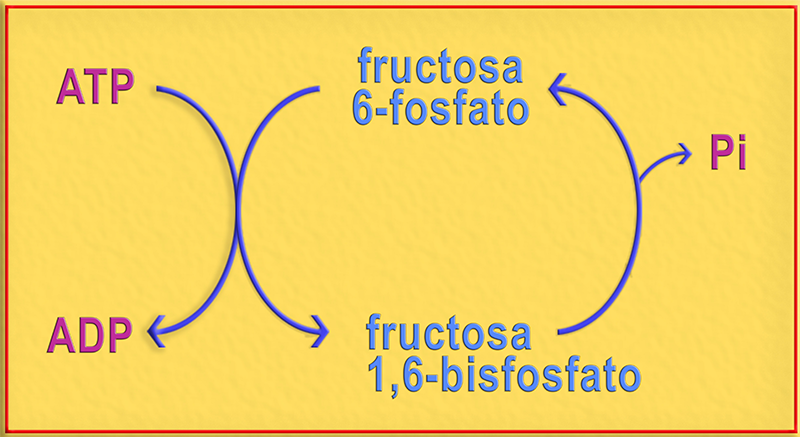
● Se trata de un ciclo de sustrato, proceso que consiste en un par de reacciones tales que el sustrato de una enzima es el producto de otra y viceversa.
El esquema muestra la fosforilación de la fructosa 6-fosfato acoplada a la hidrólisis del ATP, formando fructosa 1,6-bisfosfato, así como la pérdida posterior de un fosfato inorgánico (Pi) para formar nuevamente la fructosa 6-fosfato. Se observa que tiene lugar la hidrólisis del ATP sin producir trabajo metabólico.
● En relación con el significado metabólico, actualmente se interpreta que los ciclos de sustrato (antes llamados “ciclos fútiles”) pueden servir para amplificar determinadas señales metabólicas, o bien, para generar calor producido por la hidrólisis del ATP.
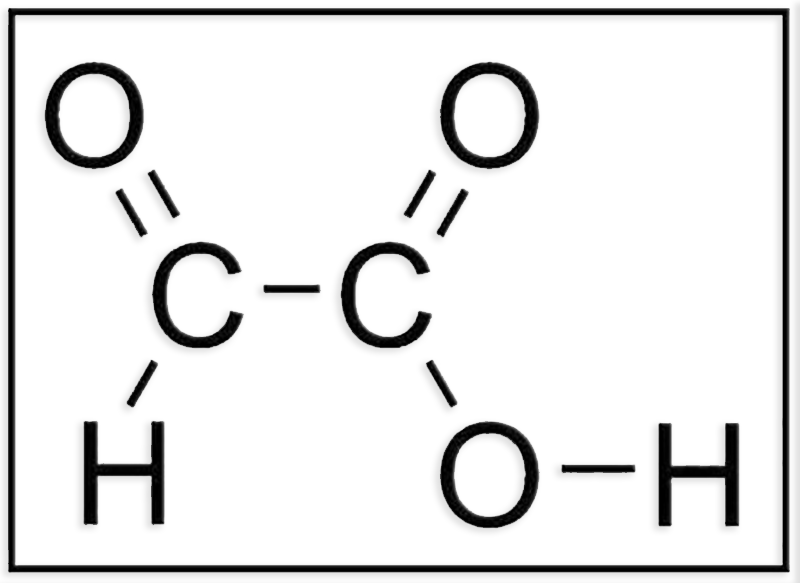
2.- Identifique el modelo molecular adjunto (C, negro. O, azul. H, blanco). Escriba la fórmula desarrollada. En relación con las células vegetales, ¿qué función desempeña?

● Se trata del ácido glioxílico (CHO―COOH). Su forma ionizada se designa como glioxilato (CHO―COO-).
● La fórmula desarrollada es:
● El ciclo de glioxilato permite la conversión en glucosa de la grasa almacenada en la semilla. Durante la germinación, las plantas realizan la conversión neta de dos moléculas de acetil-CoA en oxalacetato y por tanto en glucosa, ya que es también un intermediario de la gluconeogénesis.
Este ciclo metabólico tiene lugar en los glioxisomas y permite a las semillas en germinación convertir los triglicéridos de reserva en glucosa, puesto que los ácidos grasos que contienen se oxidan a acetil-CoA. Esto sucede hasta que la planta empieza a sintetizar glucosa a partir de la fotosíntesis.
3.- Interprete el esquema adjunto.
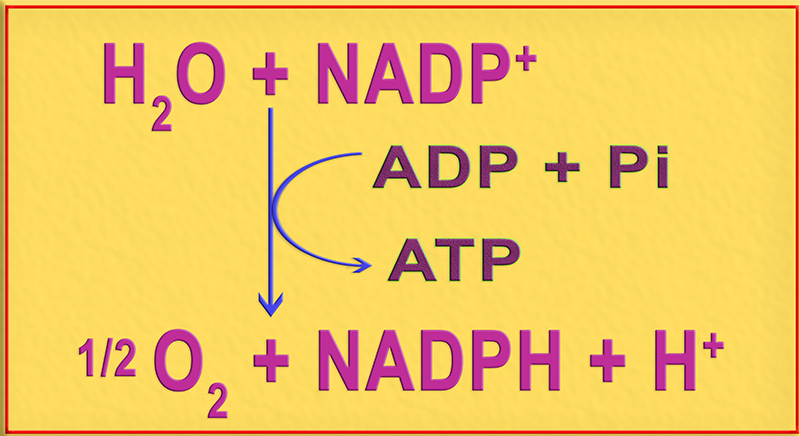
Se trata del resumen de la fase luminosa (fotoquímica) de la fotosíntesis oxigénica, teniendo lugar tres acontecimientos: fotólisis del agua, fotorreducción y fotofosforilación. Dado que el O2 se libera como subproducto, el resultado neto es: ATP y poder reductor (NADPH).
Así, pues, la energía luminosa captada por los pigmentos fotosintéticos del cloroplasto, localizados en las membranas de los tilacoides, sirve para reducir la coenzima NADP+, utilizando agua como donador de electrones, así como para formar ATP (fotofosforilación).
4.- ¿En qué se transforma la ribulosa 1,5-difosfato?
Hay que tener en cuenta que la enzima RuBisCO, o sea, ribulosa bisfosfato carboxilasa-oxigenasa (u oxidasa), es bifuncional y puede actuar sobre el sustrato como carboxilasa o como oxigenasa, en función de las cantidades relativas de CO2 y de O2.
● Carboxilación. La ribulosa 1,5-difosfato (RuDP) lleva a cabo la fijación de CO2 y se escinde originando dos moléculas de 3 carbonos: fosfoglicerato. Es la primera reacción del ciclo de Calvin (fase oscura de la fotosíntesis):
• RuDP + CO2 + H2O ―> 2 fosfoglicerato
● Oxigenación. Cuando la concentración de O2 es relativamente alta, la RuDP se rompe y origina dos moléculas: una de 3 carbonos y otra de 2. La reacción es:
• RuDP + O2 ―> fosfoglicerato + fosfoglicolato
Las transformaciones de este ácido de 2 carbonos (fosfoglicolato) constituyen la vía metabólica de la fotorrespiración, proceso que resta eficacia al fotosintético y que tiene lugar en tres orgánulos: cloroplastos, peroxisomas y mitocondrias.
5.- Identifique el modelo molecular adjunto (C, negro. O, azul. H, blanco. Grupo fosfato, rojo). ¿En qué vías metabólicas interviene?
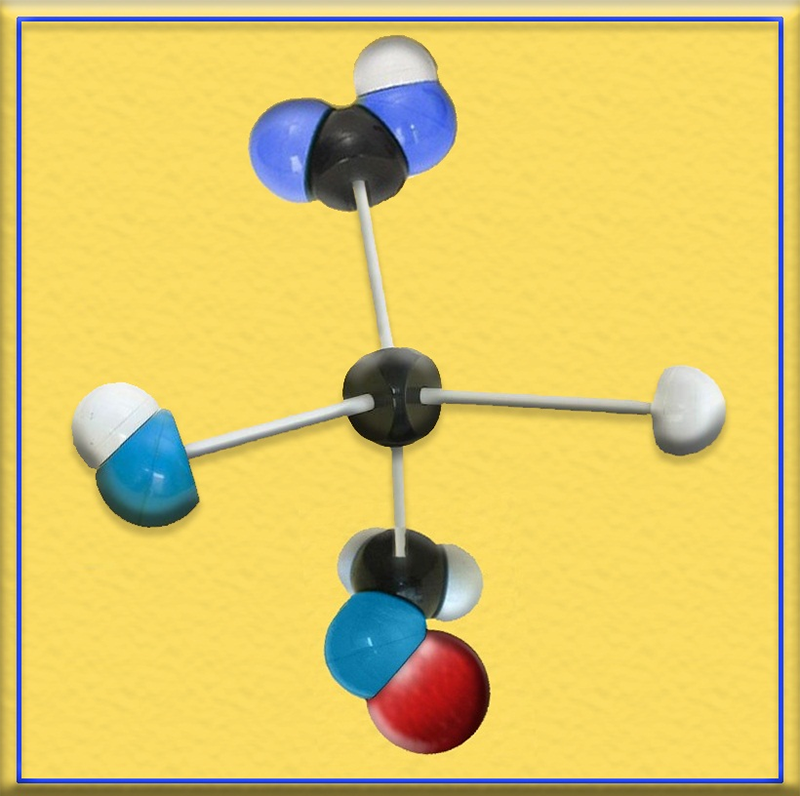
● Se trata del ácido 3-fosfoglicérico.
● El 3-fosfoglicerato interviene principalmente en la glucolisis, la gluconeogénesis y el ciclo de Calvin (fase oscura de la fotosíntesis).
6.- ¿Cuál es la finalidad metabólica de la vía de las pentosas fosfato?
La llamada vía de las pentosas es una ruta secundaria para el metabolismo de la glucosa y tiene tres funciones principales:
● Generar potencial reductor en forma de NADPH.
● Sintetizar pentosas (D-ribosa 5-fosfato especialmente)
● Incorporar los excedentes de pentosas fosfato procedentes de la dieta en el metabolismo de la glucosa.
7.- ¿En qué vía metabólica interviene la fosfoglucomutasa y qué función realiza?
● La enzima fosfoglucomutasa participa tanto en la glucogenolisis como en la glucogenogénesis.
● La fosfoglucomutasa es una enzima isomerasa que cataliza la conversión de la glucosa 1-fosfato, liberada en la degradación del glucógeno, en glucosa 6-fosfato, metabolito intermediario en la glucolisis y en la vía de las pentosas.
De modo reversible, la fosfoglucomutasa cataliza la conversión de la glucosa 6-fosfato en glucosa 1-fosfato para su activación e incorporación al glucógeno.
8.- Indique la función de las enzimas responsables de la glucogenolisis.
● La enzima glucógeno fosforilasa rompe los enlaces α (1―>4) entre las glucosas lineales y las libera en forma de glucosa 1-fosfato. Esto supone un ahorro energético ya que basta isomerizar la glucosa 1-fosfato a glucosa 6-fosfato para metabolizarla.
● La llamada enzima desramificante del glucógeno tiene actividad α (1―>6) glucosidasa, liberando la glucosa que se encuentra en los puntos de ramificación en forma de glucosa libre.
9.- La gluconeogénesis es un proceso especialmente activo en los rumiantes. Proponga una explicación.
El estómago de los rumiantes se comporta como una cámara de fermentación en la que la celulosa se hidroliza mediante las celulasas de bacterias y de otros microbios celulolíticos, resultando abundante glucosa, que fermenta y produce lactato, propionato, acetato y butirato. El lactato y el propionato, una vez absorbidos, son metabolizados formando glucosa en el hígado (gluconeogénesis).
10.- En relación con el glucógeno, ¿qué función desempeña la UDP-glucosa?
● La UDP-glucosa (uridina difosfato glucosa) se forma en la siguiente reacción:
UTP + glucosa 1-fosfato ―> UDP-glucosa + PPi
El PPi es hidrolizado por una pirofosfatasa resultando 2 fosfatos (2 Pi).
● La función de la UDP-glucosa es actuar como dador de restos de glucosa, que son transferidos por la glucógeno sintasa a la cadena de glucógeno que está en formación (glucogenogénesis).
La reacción es:
UDP-glucosa + (glucosa)n ―> UDP + (glucosa)n+1
11.- ¿Qué relación hay entre la glucogenina y la glucogenogénesis?
La glucogenina es una glucoproteína que, gracias a los residuos de tirosina (aminoácido con un grupo OH terminal en la cadena lateral), actúa como núcleo o anclaje para iniciar la molécula de glucógeno, de forma que el primer enlace que se establece es:
(Tir)-OH + HO-(glucosa) —> (Tir)-O-(glucosa) + H2O.
A partir de este momento se van adicionando glucosas mediante la acción catalítica de la enzima glucógeno sintasa, que las une con enlace glucosídico alfa 1—>4.
Por consiguiente, la glucogenina es necesaria para iniciar la síntesis del glucógeno, proceso llamado glucogenogénesis, que requiere la presencia de glucosa activada en forma de UDP-glucosa.
12.- ¿Pueden los mamíferos sintetizar glucosa a partir de los ácidos grasos? Justifique la respuesta.
Los ácidos grasos de número par de carbonos no son gluconeogénicos, es decir, no son capaces de producir glucosa, ya que al metabolizarse por β-oxidación generan en la última etapa dos moléculas de acetil-CoA, metabolito que no puede entrar en la gluconeogénesis.
Sin embargo, los ácidos grasos de número impar de carbonos, aunque escasos en la dieta, al final de su oxidación producen un resto de propionil-CoA capaz de metabolizarse en succinil-CoA y posteriormente en oxalacetato, que es un metabolito intermediario de la gluconeogénesis.
Esquema:
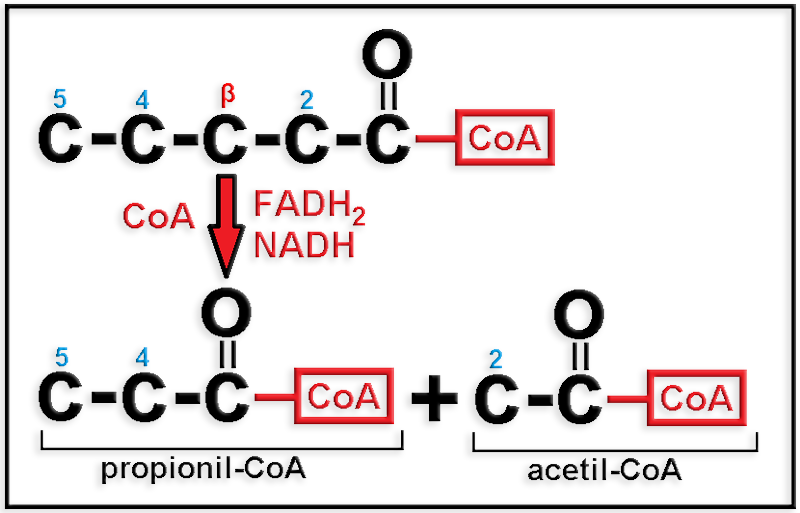
En conclusión: son gluconeogénicos los ácidos grasos de número impar de carbonos, pero no los de número par.
13.- ¿Cuál es el nombre del compuesto representado? ¿Cómo se origina? ¿Cómo se metaboliza?
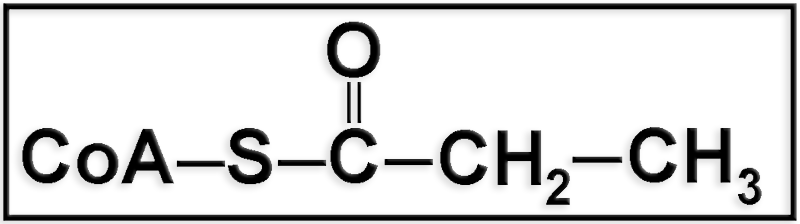
● Se trata del propionil-CoA.
● Se origina en la β-oxidación de los ácidos grasos con número impar de átomos de carbono, pues el acil-CoA se va acortando sucesivamente en dos carbonos, de modo que cuando quedan sólo cinco, o sea, [5C]-CoA, se parte en un fragmento de dos, [2C]-CoA, y otro de tres, [3C]-CoA, es decir, acetil-CoA y propionil-CoA, respectivamente.
El propionil-CoA también puede provenir del metabolismo de las cadenas carbonadas de algunos aminoácidos.
● El resto de propionil-CoA se puede metabolizar y transformar en succinil-CoA (metabolito intermediario del ciclo de Krebs), que posteriormente se convierte en oxalacetato, compuesto que puede derivar hacia la gluconeogénesis.
14.- Suponga que un grupo de cobayas, en el laboratorio, es alimentado desde el nacimiento con una dieta carente de grasa. Indique si se presentará alguna deficiencia en colesterol, fosfoacilglicéridos o prostaglandinas.
● El colesterol se puede sintetizar a partir del acetil-CoA.
● Los fosfoacilglicéridos provienen del ácido fosfatídico, una molécula que se forma a partir de acetil-CoA y de glicerol 3-fosfato.
● Las prostaglandinas derivan fundamentalmente del araquidónico, que es un ácido graso esencial (debe ser incorporado en la dieta). Por esta razón, una dieta libre de grasa provocaría una deficiencia de prostaglandinas.
15.- Identifique la siguiente representación molecular. ¿Cómo se origina? ¿Cómo puede metabolizarse?
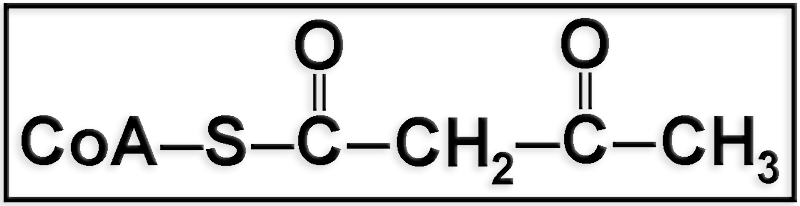
● Se trata del acetoacetil-CoA.
● Se origina mediante una reacción de condensación entre dos moléculas de acetil-CoA.
● El acetoacetil-CoA puede metabolizarse reaccionando con otra molécula de acetil-CoA para dar un compuesto intermediario tanto en la biosíntesis de colesterol como en la formación de cuerpos cetónicos.
16.- Interprete el proceso adjunto.
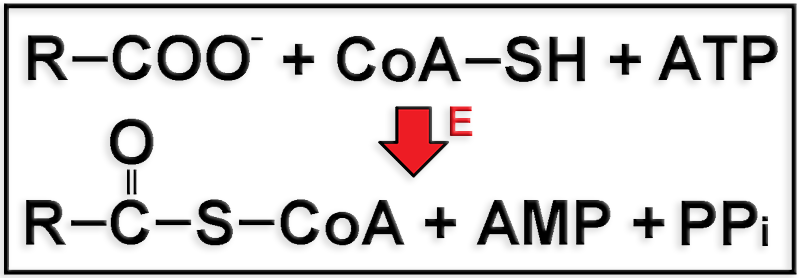
Se trata de la activación de los ácidos grasos antes de pasar a la matriz mitocondrial, mediante la acción catalítica de la enzima acil-CoA sintetasa (E), que se halla en la membrana externa de la mitocondria.
Los ácidos grasos (R-COO-) se activan uniéndose a una molécula de coenzima A (CoA-SH), con gasto de ATP, resultando acil-CoA, AMP y pirofosfato (PPi). La escisión del PPi en dos fosfatos (2 Pi) mediante una pirofosfatasa hace irreversible la reacción.
El acil-CoA resultante es transportado al interior de la mitocondria utilizando, en general, la lanzadera de la carnitina.
17.- Explique la función biológica de la carnitina.
La función principal de la carnitina es transportar a los ácidos grasos de cadena larga activados (acil-CoA) desde el citosol hasta la matriz mitocondrial o mitosol.
Los ácidos grasos de cadena más larga de 20 carbonos se oxidan en los peroxisomas, mientras que los menores de 10 carbonos entran directamente por difusión en la mitocondria. Sin embargo, los que poseen entre 10 y 20 átomos de carbono, mayoritarios en aceites y grasas de nuestra dieta, necesitan ser activados en la membrana mitocondrial externa antes de ser introducidos en la matriz.
En la activación se obtienen moléculas llamadas genéricamente acil-CoA, que no pueden pasar a través de la membrana mitocondrial interna. Para salvar este inconveniente existe un mecanismo especial de transporte mediado por la carnitina, que actúa fundamentalmente en el espacio intermembrana y cuyos pasos principales son:
• El resto acilo se transfiere desde el acil-CoA a la carnitina formando acil-carnitina.
Es decir: acil-CoA + carnitina ―> CoA + acil-carnitina
• La acil-carnitina actúa como una lanzadera a través de la membrana mitocondrial interna por medio de una translocasa.
• El grupo acilo de la acil-carnitina se transfiere entonces a una molécula de CoA de la matriz mitocondrial reconstituyendo el acil-CoA.
Es decir: acil-carnitina + CoA ―> acil-CoA + carnitina
• La carnitina libre reinicia el proceso.
18.- Cite el nombre correspondiente a la representación adjunta. ¿Qué función desempeña?
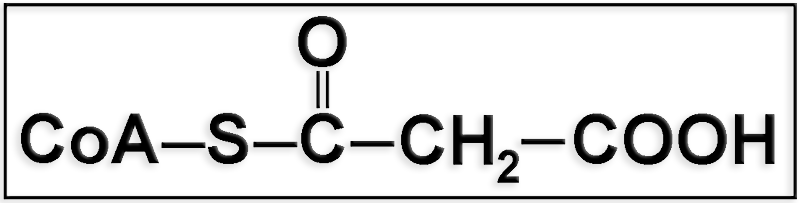
● Se trata del malonil coenzima A (malonil-CoA).
● Aunque el malonil-CoA desempeña su función principal como precursor de la síntesis de ácidos grasos, conviene reseñar que, indirectamente, tiene un papel regulador en la β-oxidación puesto que inhibe el funcionamiento del sistema lanzadera de la carnitina, frenando de esta manera dicho proceso.
19.- ¿En qué consiste la β-oxidación de los ácidos grasos? Haga un esquema del proceso partiendo de un ácido graso saturado con 10 átomos de carbono. ¿Cuál sería el resultado?
● La β-oxidación consiste en la degradación de los ácidos grasos mediante un proceso por etapas, en cada una de las cuales se forma una molécula de acetil CoA y dos de coenzimas reducidos (FADH2 y NADH).
El ácido graso propuesto en el enunciado es: CH3―(CH2)8―COOH.
En cada etapa del proceso el ácido graso se acorta en dos carbonos generando: acetil-CoA, FADH2 y NADH. Como inicialmente posee 10 carbonos, el ciclo de oxidación debe repetirse 4 veces.
● Esquema:
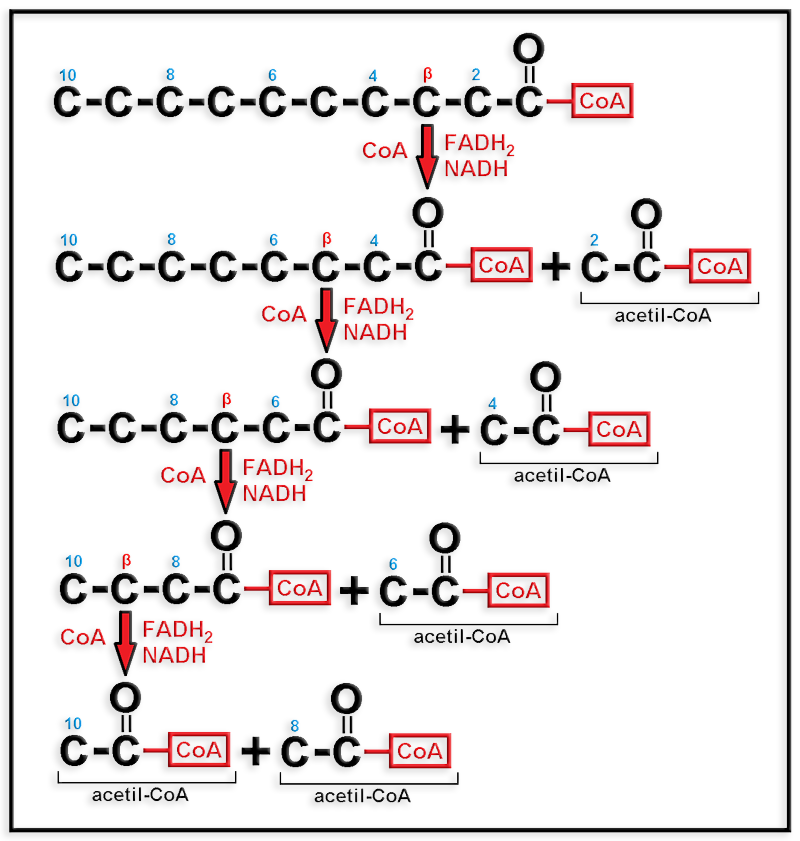
● Resultado: 5 moléculas de acetil-CoA, 4 de FADH2 y otras 4 de NADH.
20.- Indique el resultado de la degradación (β-oxidación) del ácido palmítico y calcule cuánto ATP se produce.
El ácido palmítico es un ácido graso saturado que tiene 16 carbonos.
En el proceso de la β-oxidación, cada acil-CoA se degrada mediante una secuencia repetitiva de cuatro reacciones enzimáticas, de forma que cada vez que el ácido se acorta en dos carbonos se va generando: acetil-CoA, NADH y FADH2.
Como el palmítico (palmitato) tiene 16 C son necesarios 7 ciclos de β-oxidación, dando como resultado: 8 moléculas de acetil-CoA, 7 de NADH y 7 de FADH2.
Para calcular el balance energético debe tenerse en cuenta que en la formación del acil-CoA (activación) se consume 1 ATP (ver nota 1). Además hay que contabilizar el ATP generado en la fosforilación oxidativa mediante la cadena respiratoria (CR): si el donador de electrones es la coenzima FADH2 se forman 2 ATP, y si es la NADH, 3 ATP (ver nota 2).
Por consiguiente:
● 8 moléculas de acetil-CoA que entran en el ciclo de Krebs:
• 8 x 1 GTP (ATP) = 8 ATP.
• 8 x 3 NADH (CR) ―> 24 x 3 ATP = 72 ATP.
• 8 x 1 FADH2 (CR) ―> 8 x 2 ATP = 16 ATP.
● 7 etapas de la β-oxidación:
• 7 NADH (CR) ―> 7 x 3 ATP = 21 ATP.
• 7 FADH2 (CR) ―> 7 x 2 ATP = 14 ATP.
Total: 131 ATP – 1 ATP (activación) = 130 ATP.
Nota 1.- Algunos autores restan 2 ATP, ya que en la activación el ATP se hidroliza dando AMP y pirofosfato (ATP ―> AMP + PPi), que se escinde en dos fosfatos (PPi ―> 2 Pi).
Nota 2.- Aunque en la mayoría de los libros de 2º de Bachillerato aparecen esos valores, la estimación que parece más correcta es que cada par de electrones cedidos por la NADH proporcionan en su transporte energía suficiente para la formación de 2,5 moléculas de ATP y 1,5 moléculas de ATP si el donador de electrones es la FADH2.
21.- Calcular el ATP que se produce en la β-oxidación del ácido esteárico.
Se puede efectuar el cálculo del ATP estableciendo las siguientes consideraciones:
● El esteárico es un ácido graso saturado que posee 18 carbonos.
● En la formación del estearil-CoA (activación) se gasta ATP, que se hidroliza en AMP y pirofosfato.
● La oxidación del estearil-CoA requiere 8 etapas de la β-oxidación, resultando: 9 acetil-CoA, 8 FADH2 y 8 NADH.
● La oxidación de cada acetil-CoA en el ciclo de Krebs produce 12 ATP.
● La oxidación de cada NADH en la cadena respiratoria (CR) produce 3 ATP, mientras que la de FADH2 da lugar a 2 ATP (ver nota).
● Por consiguiente:
• 9 acetil-CoA ―> 9 x 12 ATP = 108 ATP.
• 8 NADH (CR) ―> 8 x 3 ATP = 24 ATP.
• 8 FADH2 (CR) ―> 8 x 2 ATP = 16 ATP.
● La suma es: 148 ATP.
● Si ahora consideramos que en la activación se gastan 2 enlaces “de alta energía” (ATP ―> AMP + PPi), el rendimiento energético de la oxidación completa del estearato es de 146 ATP.
Nota.- La estimación que parece más correcta es que cada par de electrones cedidos por la NADH proporcionan en su transporte energía suficiente para la formación de 2,5 moléculas de ATP y 1,5 moléculas de ATP si el donador de electrones es la FADH2. Estos valores suelen redondearse a 3 ATP y 2 ATP, respectivamente.
22.- Se detecta la presencia de etanol en al análisis sanguíneo de un paciente que asegura no haber ingerido ningún tipo de bebida alcohólica durante mucho tiempo. Proponga una explicación.
Algunos microorganismos disponen de las enzimas necesarias para transformar el piruvato en acetaldehído y su posterior conversión en etanol.
La piruvato descarboxilasa cataliza la transformación del piruvato en acetaldehído (etanal). Esta enzima está ausente en todos los tejidos de nuestro organismo.
La enzima alcohol deshidrogenasa cataliza la conversión del etanal en etanol.
Esquema:

Así, pues, un exceso de determinada flora microbiana puede ser la causa originaria del etanol, que tras la absorción intestinal se detectaría en el plasma sanguíneo del paciente.
23.- ¿Cómo se metaboliza el etanol de las bebidas alcohólicas?
La mayor parte del alcohol, un 90-95 %, se metaboliza a nivel hepático, eliminándose la fracción restante por la vía respiratoria. Este etanol presente en el aire espirado puede ser detectado mediante un alcoholímetro, lo cual permite una estimación indirecta del grado de alcoholemia.
En el hígado, la enzima alcohol deshidrogenasa transforma el etanol en acetaldehído (etanal), y la aldehído deshidrogenasa, lo convierte en acético (acetato).
Esquema:

El acético (acetato) sigue metabolizándose y se transforma en acetil coenzima A (acetil-CoA), que puede participar en el ciclo de Krebs o en la síntesis de cuerpos cetónicos y de ácidos grasos.
24.- Indique la función de MEOS (Microsomal Ethanol-Oxidizing System).
El etanol se puede transformar en etanal (acetaldehído) mediante la vía oxidativa de un sistema de microsomas localizado principalmente en las células del hígado.
El esquema de esta variante metabólica es:
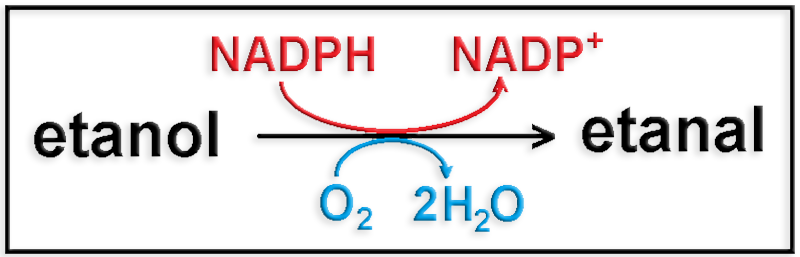
O bien: CH3―CH2OH + NADPH + H+ + O2 ―> CH3―CHO + NADP+ + 2 H2O
25.- En relación con el etanol, indique la acción de la catalasa.
Una tercera vía de degradación del etanol es mediante la catalasa, enzima presente en los peroxisomas de las células de diversos tejidos, dando como resultado la formación de acetaldehído (etanal).
Esquema:
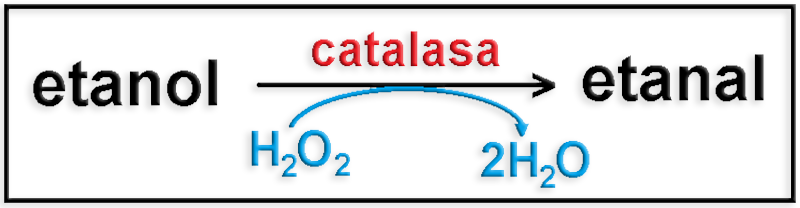
O bien: CH3―CH2OH + H2O2 ―> CH3―CHO + 2 H2O
Nota.- Existe una 4ª variante sobre el metabolismo del etanol relacionada con la formación de ésteres etílicos.
26.- ¿Por qué queda afectado el hígado por el consumo continuado o abusivo de bebidas alcohólicas? ¿Existe alguna relación con el cáncer?
El etanol de las bebidas alcohólicas (CH3―CH2OH) es oxidado a acetaldehído o etanal (CH3―CHO), que luego se convierte en acético (CH3―COOH). El ácido acético es utilizado para formar acetil-CoA, que tiene diversos destinos metabólicos.
Las reacciones son:
• CH3―CH2OH + NAD+ ―> CH3―CHO + NADH + H+
• CH3―CHO + NAD+ ―> CH3―COOH + NADH + H+
• CH3―COOH + HS-CoA + ATP ―> CH3―CO―S-CoA + ADP
● Es importante observar que el consumo de etanol provoca una gran concentración de NADH, con el consiguiente déficit de NAD+, lo cual provoca, por un lado, la inhibición de vías metabólicas tales como la oxidación de ácidos grasos y el ciclo de Krebs, y por otro, la activación de la síntesis de ácidos grasos. Al cabo del tiempo, las células hepáticas, cargadas de grasa, se vuelven disfuncionales y originan un proceso inflamatorio, la hepatitis alcohólica, enfermedad que si no se corrige conduce hacia la cirrosis hepática, situación en la que dicho órgano no puede realizar sus funciones y que tiene un pronóstico infausto.
● Por otra parte, el consumo abusivo de etanol puede aumentar el riesgo de desarrollar un cáncer debido a la toxicidad del acetaldehído, metabolito que tiene diversos efectos perjudiciales en el organismo, entre los cuales se encuentra la interferencia con el proceso de reparación del ADN defectuoso.
27.- Identifique el modelo molecular adjunto (C, negro. O, azul. H, blanco). Escriba la fórmula a pH fisiológico. ¿Cuál es su función metabólica?
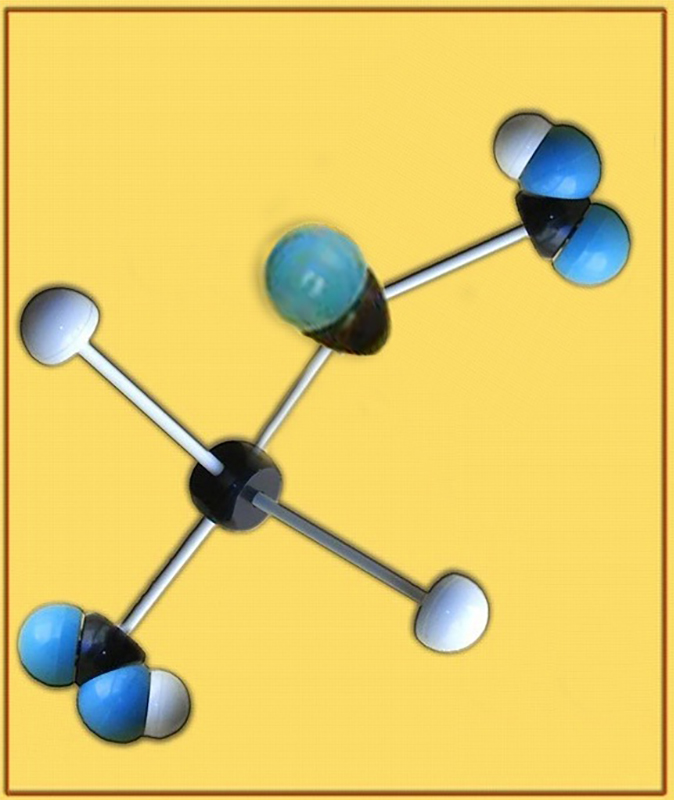
● Se trata del ácido oxalacético.
● La fórmula a pH fisiológico es:
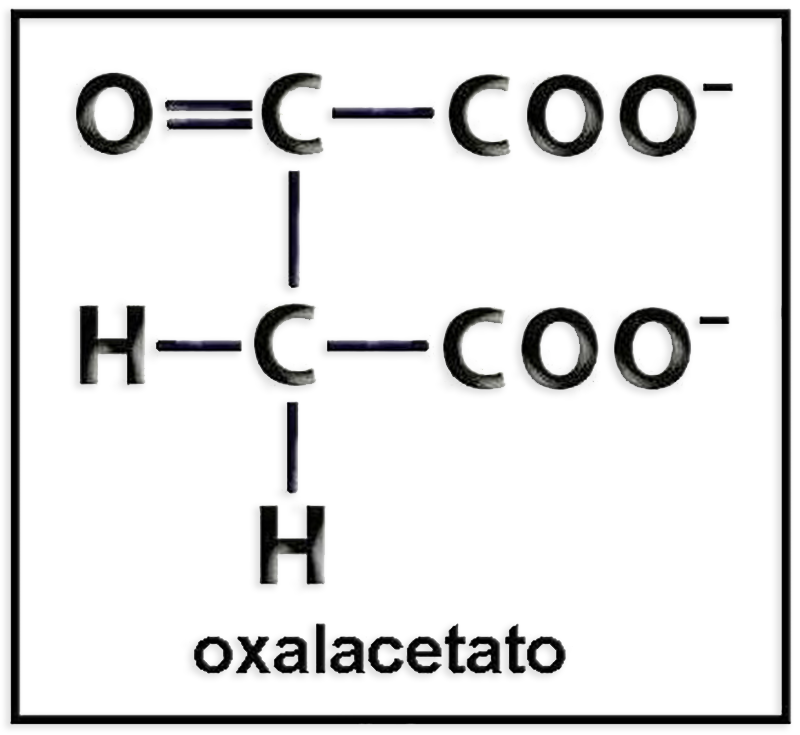
● La función metabólica del oxalacetato es diversa:
• Participar en el ciclo de Krebs (primera y última reacción).
• Intervenir en la gluconeogénesis y producir glucosa.
• Por transaminación forma aspartato, que se incorpora al ciclo de la urea.
• En las células vegetales, considerando la vía fotosintética del C4, el oxalacetato es la primera molécula que se obtiene tras la fijación del CO2 por parte del fosfoenolpiruvato.
28.- En relación con el ciclo de los ácidos tricarboxílicos, complete la siguiente reacción y añada un breve comentario.
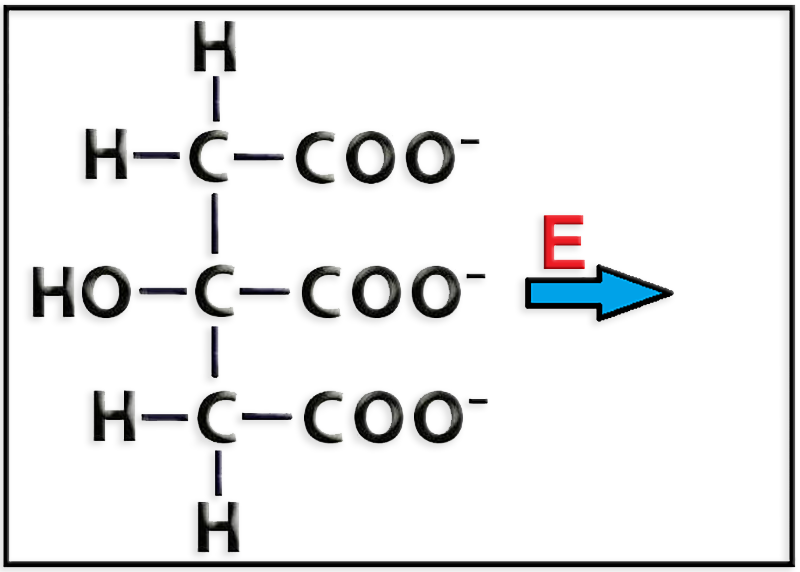
● La reacción es:
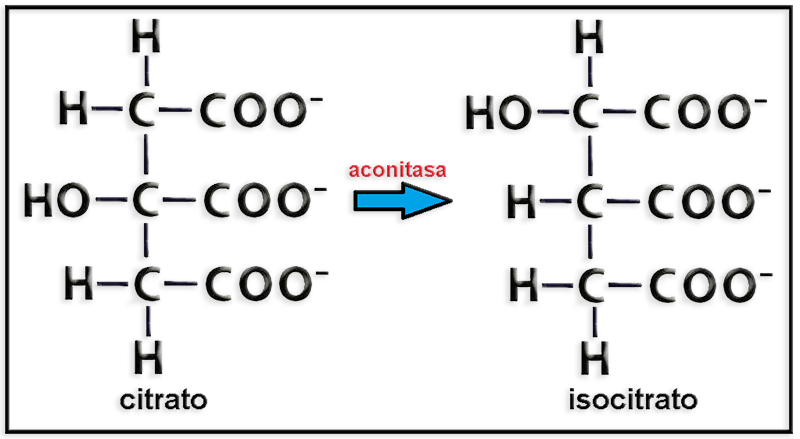
● En relación con el ciclo de Krebs, el citrato se transforma en isocitrato en una reacción catalizada por la enzima aconitasa, en la que tiene lugar la salida y la posterior entrada de una molécula de agua así como la formación de un compuesto intermedio no representado (aconitato).
29.- Identifique la representación molecular adjunta. En relación con el ciclo de Krebs, ¿cómo se origina y en qué se transforma?
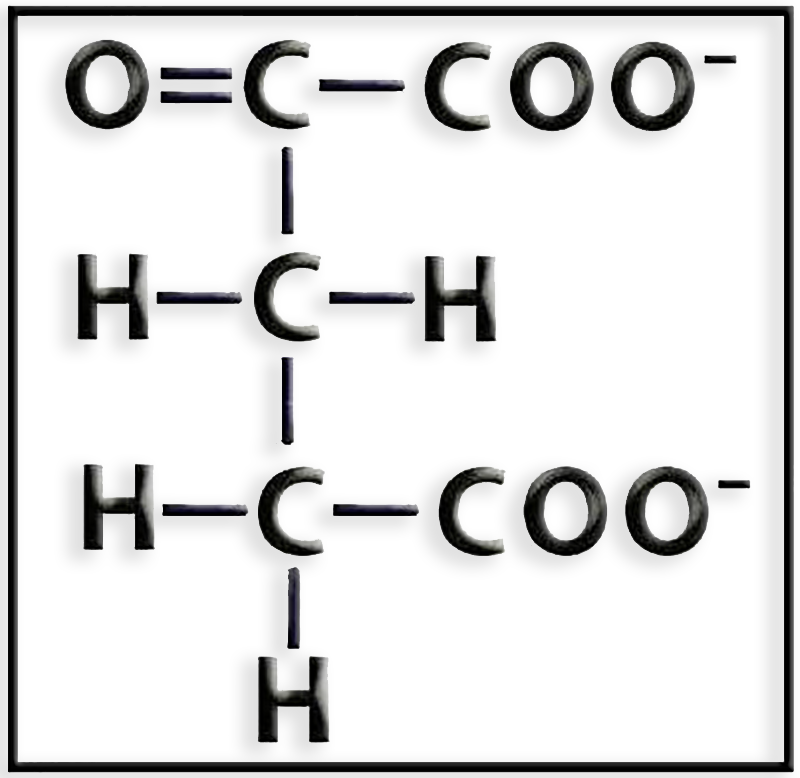
● Se trata del α-cetoglutarato.
● Se origina por descarboxilación oxidativa del isocitrato, reacción catalizada la enzima isocitrato deshidrogenasa.
La reacción es:
isocitrato + NAD+ ―> cetoglutarato + CO2 + NADH + H+
● La enzima cetoglutarato deshidrogenasa cataliza la descarboxilación oxidativa del cetoglutarato, que se transforma en succinil-CoA previa incorporación de la coenzima A.
La reacción es:
cetoglutarato + CoA-SH + NAD+ ―> succinil-CoA + CO2 + NADH + H+
30.- Identifique el modelo molecular adjunto (C, negro. O, azul. H, blanco). Escriba la fórmula a pH fisiológico. En relación con la fotosíntesis, ¿qué función desempeña?
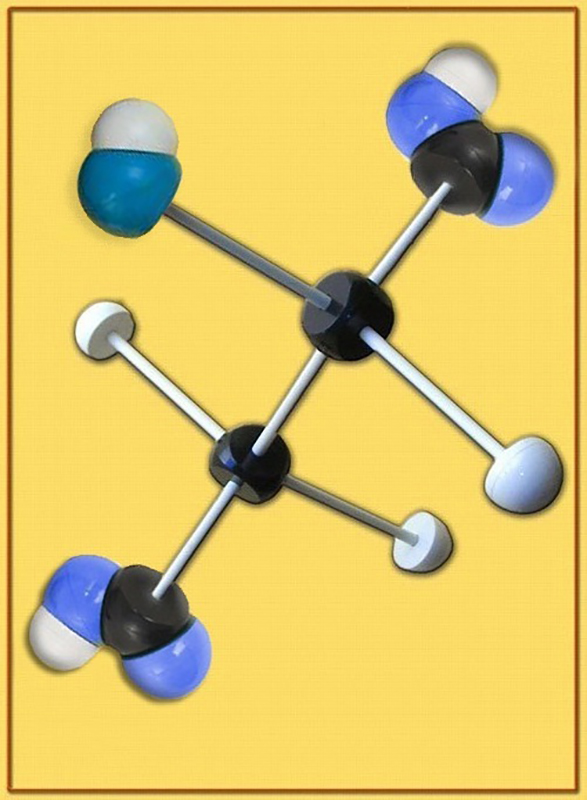
● Se trata del ácido málico (malato).
● La fórmula a pH fisiológico (o neutro) es:
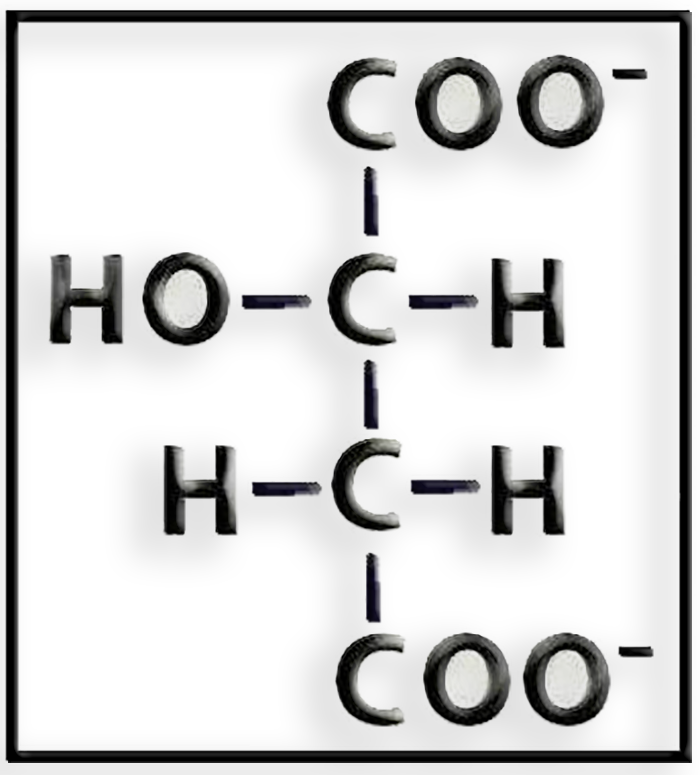
● En la vía fotosintética del C4, también llamada ruta de Hatch y Slack (sus descubridores), el malato se forma por reducción del oxalacetato en el citosol de las células del mesófilo, pasando a través de los plasmodesmos hacia las células que rodean a los vasos conductores, en cuyos cloroplastos sufre una descarboxilación oxidativa y se convierte en piruvato.
Las reacciones son:
• Citosol: oxalacetato + NADPH + H+ ―> malato + NADP+
• Cloroplastos: malato + NADP+ ―> CO2 + piruvato + NADPH + H+
El CO2 que se desprende es fijado en la ribulosa difosfato gracias a la acción catalítica de la enzima ribulosa bisfosfato carboxilasa (rubisco), activando el ciclo de Calvin.
El piruvato sale al citosol y, con gasto de ATP, forma fosfoenolpiruvato, que tras la fijación de CO2 y la pérdida de fosfato se convierte en oxalacetato.