2.5.3. Digestión celular. Orgánulos implicados.
2.5.3. Digestión celular. Orgánulos implicados.
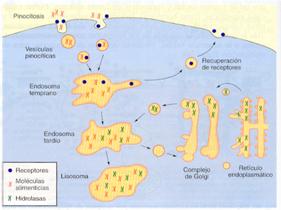
Las vesículas pinocíticas son inicialmente descargadas en unas vesículas intracelulares pequeñas e irregulares llamadas endosomas tempranos. En ellas, las moléculas endocitadas son seleccionadas y, algunas, como los receptores, pueden regresar a la membrana plasmática y reciclarse, mientras que la mayoría se dirigen a los llamados endosomas tardíos y, posteriormente, a los lisosomas. La relación entre estos tres tipos de vesículas membranosas no está totalmente aclarada. Una hipótesis es que los endosomas tempranos se desplazan lentamente hacia el interior de la célula pasando a ser endosomas tardíos. Éstos posteriormente se convertirán en lisosomas, como resultado de su fusión con pequeñas vesículas que transportan hidrolasas desde el complejo de Golgi y de un incremento de la acidificación.
En los lisosomas tiene lugar, finalmente, la digestión intracelular. Estos orgánulos contienen hidrolasas lisosomales, que previamente se habrán formado en los ribosomas del retículo endoplasmático rugoso, del que pasan al complejo de Golgi, donde son seleccionadas y empaquetadas con destino a los lisosomas.
En el medio ácido del lisosoma las hidrolasas descomponen las partículas y macromoléculas alimenticias liberando moléculas pequeñas como aminoácidos, monosacáridos, nucleótidos, etc. Los productos finales de la digestión salen del lisosoma mediante transportadores específicos de la membrana del lisosoma y, posteriormente, se pueden incorporar a diversos procesos metabólicos de la célula o ser excretados.
|
|
|
Vía endocítica, desde la membrana plasmática a los lisosomas. |
Aparte de la digestión de vesículas pinocíticas, las células pueden degradar partes envejecidas u obsoletas de la propia célula mediante un proceso de autofagia (formándose autofagolisosomas).
Una tercera vía tiene lugar en los protozoos y células especializadas en realizar la fagocitosis. En éstas, los lisosomas se unen a los fagosomas (formando fagolisosomas), para digerir los microorganismos y grandes partículas ingeridas.
2.5.4. Excreción: exocitosis.
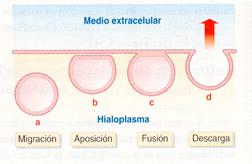
El fenómeno de expulsión de sustancias por mediación de vesículas y, por tanto, contrario a la endocitosis, se denomina exocitosis . Precisamente, lo más sencillo es concebirlo como un proceso inverso al de endocitosis: las vesículas que engloban las sustancias que han de ser expulsadas migran hacia la membrana plasmática; se acercan a ella y se adhieren a la misma en un fenómeno conocido como aposición; a continuación, las estructuras membranales de vesícula y membrana plasmática se, funden y, finalmente, se produce la descarga del material.
|
|
|
Fases del proceso de exocitosis: a) Migración, b) Aposición, c) Fusión y d) Descarga. |
2.5.5. Metabolismo.
Una célula es un complejo de laboratorios en el que continua y simultáneamente se producen y se destruyen las más diversas sustancias. Sus fábricas se surten de combustibles, los nutrientes, de los que extraen la energía necesaria para que todo funcione y para construir y reparar los materiales de los que está hecha la propia célula.
El complejo de fabricación es un todo unitario que funciona con el mínimo gasto. La economía exige regulación y una máxima coordinación. Cada molécula cumple su función y todas están integradas en un conjunto autorregulado: el metabolismo .
2.5.5.1. Concepto de metabolismo, catabolismo y anabolismo.
El metabolismo es el conjunto de reacciones químicas que se producen en el interior de las células y que conducen a la transformación de unas biomoléculas en otras. Las distintas reacciones químicas del metabolismo se denominan vías metabólicas y las moléculas que en ellas intervienen se llaman metabolitos.
Todas las reacciones del metabolismo están reguladas por enzimas, que son específicas para cada metabolito inicial o sustrato y para cada tipo de transformación. Las sustancias finales de una vía metabólica se denominan productos. Las conexiones existentes entre diferentes vías metabólicas reciben el nombre de metabolismo intermediario.
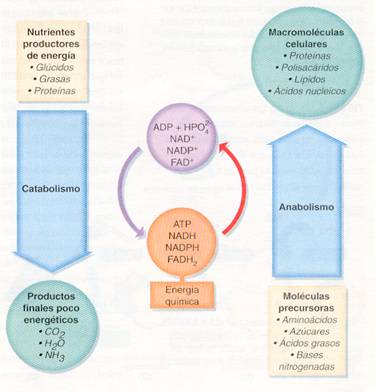
Se pueden considerar dos fases en el metabolismo: una de degradación de materia orgánica o catabolismo y otra de construcción de materia orgánica o anabolismo.
|
|
|
Características de las dos fases del metabolismo. |

· El catabolismo es la transformación de moléculas orgánicas complejas en otras más sencillas, proceso en el que se libera energía que se almacena en los enlaces fosfato del ATP.
· El anabolismo es la síntesis de moléculas orgánicas complejas a partir de otras biomoléculas más sencillas, para lo cual se necesita suministrar energía, proporcionada por los enlaces fosfato del ATP. Las moléculas del ATP pueden proceder de las reacciones catabólicas, de la fotosíntesis (en las plantas y algunos microorganismos) o de la quimiosíntesis (en otros microorganismos).
2.5.5.2. Aspectos generales del metabolismo: reacciones de oxidorreducción y ATP.
El metabolismo celular se tiene que realizar en condiciones determinadas:
- A una temperatura baja, que no desnaturalice las proteínas y pierdan su función.
- En un medio acuoso, como es el medio celular.
- A un pH determinado, que no puede variar mucho sin sufrir la célula daños importantes.
- Siguiendo las leyes de la termodinámica.
El metabolismo es una actividad celular muy compleja y siempre dirigida y coordinada por sistemas multienzimáticos que permiten a la célula:
a) Obtener energía química de su entorno, sea a través de la energía luminosa procedente del sol o a partir de compuestos orgánicos.
b) Convertir el alimento en metabolitos específicos de la célula.
c) Unir moléculas sencillas (monosacáridos, aminoácidos, nucleótidos...) para formar macromoléculas como polisacáridos, proteínas o ácidos nucleicos.
d) Sintetizar y degradar moléculas especiales que la célula necesita en momentos puntuales.

Aunque son muy diversas las biomoléculas que contienen energía almacenada en sus enlaces, es el ATP (adenosín trifosfato) la molécula que interviene en todas las transacciones de energía que se llevan a cabo en las células; por ello se la califica como "moneda universal de la energía". El ATP está formado por adenina, ribosa y tres grupos fosfato, contiene enlaces de alta energía entre los grupos fosfato; al romperse dichos enlaces se libera la energía almacenada y para volver a formarse se requiere el mismo aporte energético. En la mayoría de las reacciones celulares el ATP se hidroliza a ADP, rompiéndose un solo enlace y quedando un grupo fosfato libre, que suele transferirse a otra molécula (esto se conoce como fosforilación); sólo en algunos casos se rompen los dos enlaces resultando AMP+2 grupos fosfato. El sistema ATP 1 ADP es el sistema universal de intercambio de energía en las células.
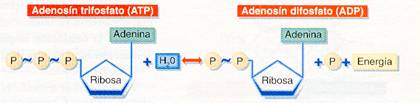
La hidrólisis de ATP suele formar ADP y un grupo fosfato libre, liberándose la energía de uno de sus enlaces.
En cualquier transformación química, solamente una parte de la energía implicada en el sistema es útil para producir trabajo; algunos la califican como energía libre ; hay otra fracción que no se puede utilizar para producir trabajo, sino que se transforma en calor, luz, cinética, etc., y que constituye la entropía.
Todas las reacciones químicas requieren una energía inicial, energía de activación, para poder romper los enlaces químicos que hay entre los átomos de los reactivos. Conviene recordar que la energía de activación es menor en las transformaciones bioquímicas debido a la acción enzimática. En unas reacciones se desprende energía y otras no se realizan si no hay un suministro de energía. Las reacciones orgánicas en las que se libera energía se conocen como exergónicas, y aquéllas en las que es necesario aportar energía para que se produzcan se denominan endergónicas. En las células ambos tipos de reacciones se realizan acopladas, de modo que en una exergónica, en la que se queman moléculas, se libera la energía que es utilizada en otra reacción endergónica que hace posible, por ejemplo, la síntesis de nuevas sustancias, el movimiento o se utiliza para mantener la temperatura corporal constante.
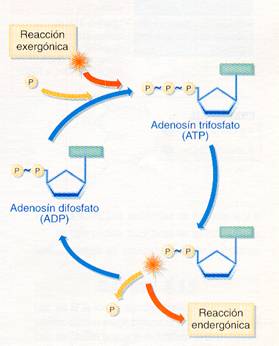
Acoplamiento de las reacciones exergónicas y endergónicas.
Las células de las plantas obtienen inicialmente la energía que necesitan a partir de energía luminosa, mientras que las animales la obtienen de los enlaces de moléculas orgánicas que ingieren y transforman. No obstante, al proceder esa materia orgánica directa o indirectamente de seres fotosintéticos, se puede decir que tanto los seres animales como los vegetales dependen de la misma fuente de energía: el sol.
Las reacciones químicas son transformaciones energéticas, en las cuales la energía almacenada en unos enlaces se transfiere a otros recién formados en moléculas diferentes; en estas reacciones los electrones pasan de un nivel energético a otro de mayor o menor energía. Con frecuencia, los electrones pasan de un átomo a otro o de una a otra molécula; estas reacciones se conocen como reacciones de oxidación-reducción , y tienen gran importancia en los sistemas biológicos.
Una oxidación es la pérdida de algún electrón; el nombre viene del oxígeno, que es un gran aceptor de electrones. Una reducción es la ganancia de algún electrón. También se puede decir que las sustancias se oxidan al ganar oxígeno o al perder átomos de hidrógeno; en ambos casos se da una pérdida de electrones. Oxidación y reducción se realizan simultáneamente, de modo que una sustancia se oxida y otra se reduce. Las reacciones de oxidación-reducción pueden afectar a un electrón en solitario, pero, con frecuencia, el electrón va unido a un protón formando un átomo de hidrógeno; en este caso la oxidación implica la pérdida de átomos de hidrógeno y la reducción la ganancia de átomos de hidrógeno. Ejemplo:
C6H12O6 + 6O2 ↔ 6CO2 + 6H2O + 686 kcal.
La glucosa se oxida, perdiendo átomos de hidrógeno que son captados por el oxígeno; los electrones pasan a un nivel energético más bajo y en la reacción se libera energía.

En los seres vivos, las reacciones que retienen energía (endergónicas), como la fotosíntesis, y aquéllas en las que se libera (exergónicas), como la glucólisis y la respiración, son reacciones de oxidación-reducción.
Existe una relación entre el grado de oxidación de un compuesto orgánico y su contenido energético: cuanto más reducido está un compuesto, mayor cantidad de energía contiene, y cuanto más oxidado se halla, menor cantidad de energía tiene.

2.5.5.3. Estrategias de obtención de energía: energía química y energía solar.
Las células siempre utilizan energía química, generalmente en forma de ATP, aunque en ocasiones emplean otros nucleótidos trifosfato, como el GTP o el UTP. Lo que diferencia unas células de otras es la fuente externa de energía que son capaces de transformar en esos nucleótidos trifosfato.
Se distinguen dos tipos de fuentes energéticas:
- La luz . Los fotones de luz contienen una cantidad de energía que depende de la longitud de onda de la luz. Las células de los organismos fototrofos disponen de sistemas para transformar la energía de los fotones en energía química, que almacenan en el ATP.
- Los compuestos químicos . Almacenan energía en sus enlaces y en su estructura. Los compuestos orgánicos suelen almacenar cantidades de energía mayores que los compuestos inorgánicos. Muchas células son capaces de obtener la energía contenida en estas moléculas mediante oxidaciones y «almacenarla» en compuestos como el ATP. Los organismos que obtienen su ATP de esta forma se denominan quimiotrofos.
|
|
|
Clasificación de las células y los organismos según su fuente de energía, y su fuente de carbono. |

2.5.5.4. Características generales del catabolismo celular: convergencia metabólica y obtención de energía.
El catabolismo comprende el metabolismo de degradación oxidativa de moléculas orgánicas, cuya finalidad es la obtención de la energía necesaria para que la célula realice sus funciones vitales.
La célula debe disponer de una última molécula a la que pueda cederle los electrones o los hidrógenos desprendidos en las rutas de oxidación. Según sea la naturaleza del aceptor de electrones, los seres vivos se pueden clasificar como aeróbicos o aerobios, si el aceptor es el oxígeno molecular (O2) o anaeróbicos o anaerobios, si es otra molécula distinta del oxigeno (NO2-, SO42-).
En términos de evolución, los procesos anaeróbicos son mucho más antiguos, pues parece ya indiscutible que las formas de vida primitiva se produjeron muchos millones de años antes de que existiera oxígeno en la atmósfera.
El catabolismo aerobio está formado por varias rutas metabólicas que conducen finalmente a la obtención de moléculas de ATP que podrán utilizarse en otros procesos que requieren aporte energético, como son las rutas del anabolismo. La energía que no se almacena se disipa en forma de calor.
La glucosa y los ácidos grasos que entran en la célula son degradados, mediante la glucólisis y la b-oxidación respectivamente, a acetil-CoA. Las proteínas se descomponen en sus aminoácidos constituyentes, formando diferentes intermediarios. Finalmente, todos ellos entran en el ciclo de Krebs y la cadena respiratoria, produciendo CO2, H2O y ATP.
Las reacciones catabólicas son convergentes , es decir, de moléculas muy diferentes se obtienen unas pocas sustancias sencillas cómo ácido láctico, urea, CO2, etc.
|
|
|
Esquema general del catabolismo aerobio en células heterótrofas. |
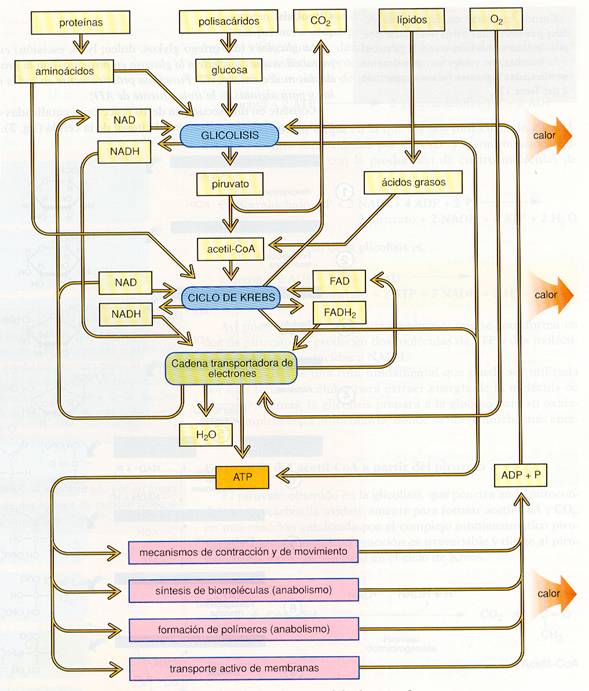
2.5.5.4.1. Glucólisis.
La glucólisis (o glicólisis) consiste en la lisis (rotura) de la glucosa en dos moléculas de piruvato (forma aniónica más frecuente del ácido pirúvico). Es un proceso que no requiere oxígeno y ocurre en el citosol de casi todas las células.
Consiste en una secuencia de diez reacciones, catalizada por otras tantas enzimas, en la que se forman diversos intermediarios con grupos fosfato. La fosforilación de todos los metabolitos excepto de la glucosa y el piruvato, impide que puedan atravesar las membranas celulares. Básicamente se pueden diferenciar dos fases:
|
|
- Fase preparatoria. Consta de cinco etapas en la que se realiza una inversión o gasto de 2 moléculas de ATP. 1. La glucosa es fosforilada a glucosa 6-fosfato con consumo de un ATP. Este proceso es irreversible en todas las células excepto en las hepáticas y las de los epitelios intestinal y renal. 2. La glucosa-6-fosfato se isomeriza a fructosa-6-fosfato. 3. Con el gasto de otro ATP, la fructosa-6-fosfato se fosforila a fructosa-l , 6-difosfato . 4. La fructosa-l,6-difosfato se hidroliza en dihidroxiacetona fosfato y gliceraldehído-3- fosfato. 5. Sólo el gliceraldehído-3-fosfato puede seguir la ruta glucolítica, pero la dihidroxiacetona se puede isomerizar a gliceraldehído-3-fosfato, por lo que a partir de esta etapa los siguientes pasos se multiplican por dos. - Fase de beneficio. Consta de otras cinco etapas en la que se recogen los beneficios de 4 moléculas de ATP que se forman por un proceso de fosforilación a partir del sustrato. 6. El gliceraldehído-3-fosfato, mediante una oxidación y fosforilación, se convierte en 1,3-difosfoglicerato. Esta etapa es básica y la primera en la que la célula obtiene energía. Parte de esta energía se conserva en el grupo fosfato del carbono 1 del 1,3-difosfoglicerato; y otra parte en el poder reductor del NADH + H+ que se genera en este paso. 7. El l,3-difosfoglicerato cede un grupo fosfato al ADP formándose ATP y 3-fosfoglicerato. 8. El grupo fosfato pasa del carbono 3 al carbono 2. 9. Por deshidratación se forma fosfoenolpiruvato con un doble enlace, y la reorganización de la molécula concentra más energía junto al grupo fosfato. 10. El fosfato es transferido al ADP, formándose otra molécula de ATP y piruvato. |
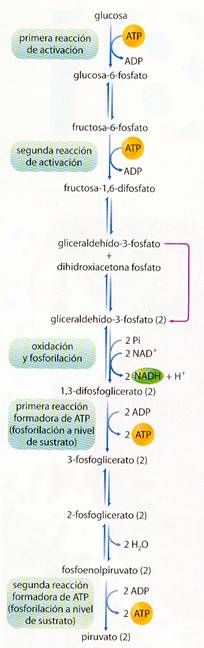
Así, en la glucólisis se consumen 2 ATP y se obtienen 4 ATP más 2 NADH + H+ , y el balance global es:
Glucosa + 2 ADP + 2 Pi + 2 NAD+ à 2 piruvato + 2 ATP + 2 (NADH + H+) + H2O
El piruvato se encuentra al final de la glucólisis en una encrucijada metabólica en la que su destino depende del tipo de célula y de la disponibilidad de oxígeno.
En condiciones anaerobias, el piruvato sigue la vía de las fermentaciones reduciéndose, con consumo de NADH + H+, a otros compuestos orgánicos como etanol o lactato.
En condiciones aerobias, el piruvato entra en las mitocondrias y es oxidado completamente hasta CO2 y H2O, a través del ciclo de Krebs y la cadena respiratoria.
Se cree que fue una de las primeras rutas metabólicas desarrolladas por los organismos primitivos para obtener energía en ausencia de oxígeno, y se ha conservado en todos los seres vivos a pesar de que, si la oxidación no continúa, su rendimiento energético es escaso: tan sólo 2 ATP.
2.5.5.4.2. Fermentación.
Las fermentaciones son procesos catabólicos en los que el aceptor final de electrones es un compuesto orgánico, generalmente formado en la propia ruta metabólica. El resultado es una oxidación incompleta del alimento. Las fermentaciones son procesos anaerobios, realizados en ausencia de oxígeno por microorganismos anaerobios estrictos o anaerobios facultativos, y, cuando no llega suficiente oxígeno, por algunas células animales o vegetales.
Los combustibles más comunes para la fermentación son azúcares, especialmente glucosa, aunque también pueden usarse otros sustratos orgánicos, como aminoácidos, ácidos orgánicos, etc. Dependiendo del producto final de la fermentación se distinguen varios tipos.
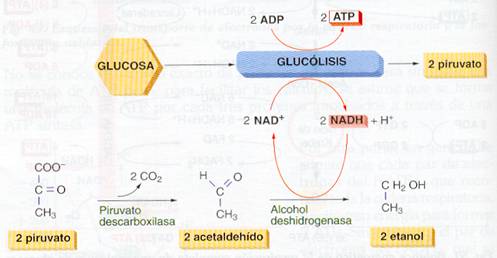
Fermentación alcohólica
Mediante la fermentación alcohólica la glucosa se transforma en 2 moléculas de etanol y 2 de CO2, produciendo 2 ATP.
Se inicia con la glucólisis que forma 2 moléculas de piruvato. El piruvato se descarboxila y pasa a acetaldehído que se reduce mediante el NADH + H+ generado en la glucólisis, formando etanol.
La fermentación alcohólica es realizada por las levaduras del género Saccharomyces, por ciertas bacterias y microorganismos, y en los tejidos de varias plantas superiores como las raíces del maíz. En la industria tiene interés lo que para los organismos fermentadores son sus productos de desecho, es decir el alcohol o el CO2. En la fabricación de bebidas alcohólicas se utilizan fundamentalmente diversas especies del género Saccharomyces que son anaerobias facultativas: S. cerevisae para fabricar cerveza, güisqui o ron; S. uvarum para el vino; S. cidrii para la sidra. En la fermentación del pan, realizada por cepas especiales de S. cerevisae, lo importante no es el etanol (que es poco y se elimina durante la cocción) sino el CO2 que esponja la masa.
|
|
|
Fermentación alcohólica. |
Fermentación láctica.
Consiste en la formación de lactato a partir de la glucosa, que a su vez puede proceder de la lactosa. En primer lugar la lactosa se hidroliza en glucosa y galactosa; y esta última se isomeriza a glucosa, con lo que el resultado son dos moléculas de glucosa. Se produce la glucólisis y se forman dos piruvatos por cada glucosa. Finalmente elpiruvato se reduce a lactato, consumiendo el NADH + H+ producido en la glucólisis. Como en el caso anterior, el rendimiento energético es de 2 ATP por cada glucosa (4 ATP a partir de lactosa).
|
|
|
Fermentación láctica. |
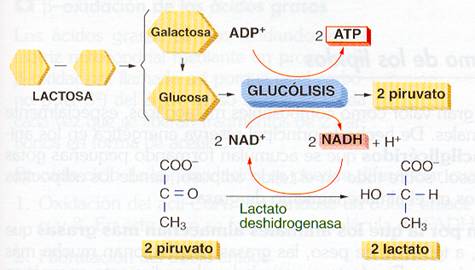
Los microorganismos que realizan la fermentación láctica son bacterias, como las de los géneros Lactobacillus, Streptococcus o Leuconostoc; y tienen importantes aplicaciones industriales para la obtención de derivados lácteos como el queso, la mantequilla, la cuajada o el yogur.
La fermentación láctica también ocurre, cuando falta oxígeno, en ciertas plantas, como las patatas y, sobre todo, en células animales, como las fibras del músculo estriado, cuando no hay suficiente oxígeno para efectuar un sobreesfuerzo físico y el ácido pirúvico procedente de la glucólisis no puede oxidarse de manera aerobia y se transforma en ácido láctico. La acumulación de ácido láctico da lugar a la formación de unos pequeños cristales que pinchan el músculo y producen los dolores conocidos como «agujetas».
Cuando el producto de la fermentación es sólo ácido láctico, se denomina fermentación homoláctica; si aparecen otros compuestos como el etanol y el dióxido de carbono, se trata de una fermentación heteroláctica.
En los dos tipos de fermentación estudiados se cumple que:
- Los productos finales son compuestos orgánicos reducidos, que pueden ser todavía oxidados y utilizados como fuente de energía. La degradación de los combustibles orgánicos mediante fermentación anaerobia no es completa.
- El aceptor de electrones del NADH + H+ es una molécula orgánica (el ácido pirúvico o el acetaldehído).
- No se obtiene más ATP en el proceso que el obtenido en el proceso de la glucólisis.
- El coenzima reducido obtenido en la glucólisis es oxidado en su totalidad en la segunda fase. Por tanto, el balance neto de coenzimas es nulo.
Otras fermentaciones, pueden ser la butírica , que consiste en la descomposición de sustancias glucídicas de origen vegetal, como el almidón y la celulosa y tiene gran importancia, ya que contribuye a la descomposición de los vegetales en el suelo.
La fermentación pútrida, llamada también putrefacción que se diferencia de las demás fermentaciones en que los sustratos que se degradan son de naturaleza proteica o aminoacídica. Los productos obtenidos en esta fermentación son orgánicos y malolientes como el indol, la cadaverina y el escatol, a los que deben el olor los cadáveres animales y restos vegetales. Algunas putrefacciones, no obstante, dan productos poco desagradables, por lo que son utilizadas para producir los sabores típicos de algunos quesos y vinos.
2.5.5.4.2.3. Respiración: ciclo de Krebs, cadena respiratoria y fosforilación oxidativa.
La respiración es el conjunto de procesos catabólicos que consisten en la degradación de sustancias orgánicas -combustible- para formar la energía que le célula necesita. Estas sustancias orgánicas pueden, ser de origen extracelular o sustancias de reserva almacenadas por la célula.
La obtención de energía puede realizarse con necesidad de oxigeno (respiración aerobia u oxibiótica), o en ausencia de dicha molécula (respiración anaerobia o anoxibiótica).
En la respiración aerobia el oxígeno molecular (O2) es el que acepta los hidrógenos para formar agua, y en la respiración anaerobia la sustancia que se reduce es diferente del oxígeno, pero en todo caso una sustancia inorgánica como el NO3-, el S04=, etc.
La energía desprendida en la respiración esta cuantificada, es decir, se desprende poco a poco en forma de moléculas de ATP, para que la célula pueda utilizarla y no le sea perjudicial.
La respiración aerobia, que tiene lugar en el citosol y en las mitocondrias, al ser degradados los combustibles de forma total (los productos finales son CO2 y H2O) el rendimiento energético es el máximo. Este tipo de respiración está generalizado a, prácticamente, todos los seres vivos.
La respiración anaerobia es mucho más ineficaz energéticamente que la anterior, pues los productos que quedan, contienen todavía energía. La presentan solamente algunas bacterias.
La respiración aerobia , la más importante, por ser la más empleada por los seres vivos, y la que más energía produce, se realiza en varias fases, que son: el ciclo de Krebs, la cadena respiratoria y la fosforilación oxidativa.
CICLO DE KREBS
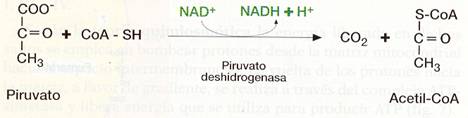
El piruvato obtenido en la glucólisis pasa, por transporte facilitado, a la matriz mitocondrial, donde se convierte en acetil-CoA mediante un proceso de oxidación y descarboxilación, en el que interviene el complejo multienzmático deshidrogenasa.
El grupo carboxilo se desprende formando CO2, y queda un grupo acetilo de carbonos que se une al CoA y se oxida, al tiempo que el NAD+ se reduce a NADH + H+.
Esta reacción es irreversible y dirige al piruvato hacia su oxidación en el ciclo del Krebs.
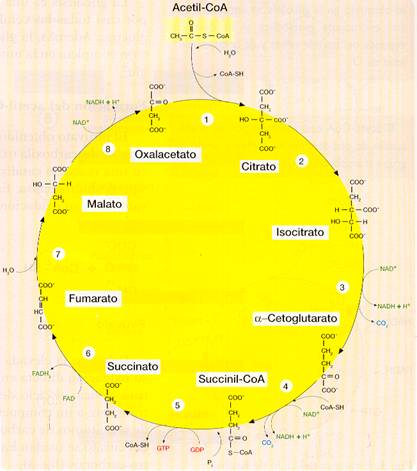
Este ciclo, consiste en una secuencia de reacciones a través de las cuales se lleva a cabo la oxidación final y total de la mayoría de los combustibles metabólicos. Se inicia con la incorporación al ciclo del acetil-CoA procedente de la glucosa (o, en otros casos, de ácidos grasos o aminoácidos). También se denomina ciclo del ácido cítrico o de los ácidos tricarboxílicos porque interviene el ácido cítrico (citrato en su forma aniónica) que posee tres grupos carboxilo (-COOH).
Este ciclo se considera el centro del metabolismo aerobio, en el que confluyen la mayoría de los procesos catabólicos e, incluso, algunas vías anabólicas (por eso se dice que es una ruta anfibólica). Tiene lugar en la matriz mitocondrial, donde se encuentran las enzimas necesarias para cada paso.
El acetil-CoA se incorpora al ciclo de Krebs y se producen 8 reacciones:
1. El grupo acetilo del acetil-CoA se condensa con el oxalacetato formando citrato, una molécula de 3 grupos carboxilos y 6 carbonos.
2. El citrato se isomeriza a isocitrato que es más fácilmente oxidable.
3. Por descarboxilación oxidativa el isocitrato se convierte en α-cetoglutarato. En esta reacción se desprende CO2 y se forma NADH + H+.
4. De nuevo tiene lugar una descarboxilación oxidativa en la que el α-cetoglutarato pasa a succinil-CoA. También aquí se desprende CO2 y se forma NADH + H+.
5. El succinil-CoA pierde la CoA y pasa a succinato, liberando la energía suficiente como para que se forme GTP mediante un proceso de fosforilación a partir del sustrato.
6. El succinato se oxida a fumarato, mediante una reacción acoplada en la que la coenzima FAD se reduce a FADH2.
7. Por hidratación del doble enlace del fumarato se forma malato.
8. A partir del malato se regenera el oxalacetato con el que se iniciaba el ciclo. La oxidación del malato se acopla a la formación de otra molécula de NADH + H+.
En resumen, la reacción global del ciclo de Krebs es la siguiente:
Acetil-CoA + 3 NAD+ + FAD + GDP + Pi + H2O —> 2 CO2 + CoA-SH + 3 (NADH + H+) + FADH2 + GTP
En cada vuelta del ciclo:
- Entra un grupo acetilo (dos átomos de carbono) que es oxidado completamente (por lo que salen del ciclo otros dos carbonos en forma de CO2).
- Tres moléculas de NAD+ son reducidas a NADH + H+.
- Una molécula de FAD es reducida a FADH2.
- Se forma una molécula de GTP equivalente al ATP.
A continuación, el NADH + H+ y el FADH2 se oxidan mediante la cadena de transporte electrónico mitocondrial generando ATP.
Esquema del ciclo de Krebs.
CADENA RESPIRATORIA Y FOSFORILACION OXIDATIVA
Aunque el ciclo de Krebs es característico de la respiración aerobia, habrás observado que en ninguna de sus reacciones interviene el oxígeno molecular. La utilización del oxígeno es exclusiva de esta última fase, en la que los electrones captados por el NADH+ H+ y el FADH2 son transportados hasta el oxígeno molecular para formar agua.
El transporte de electrones se inicia cuando una molécula de NADH + H+ o de FADH2 se oxida y cede H + y electrones a una de las, aproximadamente, 15 moléculas que forman la cadena de transporte electrónico situada en la membrana mitocondrial interna.
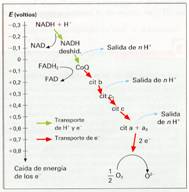
El transporte se realiza a través de una serie de reacciones de óxido-reducción. Cada reacción hace intervenir a dos constituyentes de la cadena respiratoria con potenciales redox diferentes. El potencial redox mide la afinidad de un transportador por los electrones. La afinidad es mayor cuanto mayor es el potencial redox. Los electrones se desplazan desde el constituyente que tiene potencial redox menor, hacia el que lo presenta mayor.
Las proteínas transportadoras están agrupadas en tres grandes complejos. Cada grupo posee un potencial redox más positivo que el anterior; los electrones descienden en cascada desde el NADH + H+ hacia los grandes complejos de enzimas, situados en niveles energéticos cada vez menores, hasta que finalmente son transferidos al oxígeno.
El NADH+ H+ cede H+ y electrones al complejo I, mientras que el FADH2 los cede directamente al coenzima Q, al que también van a parar los del complejo I. El coenzima Q transfiere los electrones al complejo II que, a través del citocromo-c, pasan hasta el complejo III. Finalmente, los electrones reducen el oxígeno y forman agua.
|
|
|
|
Distintas representaciones de la cadena respiratoria. |
|
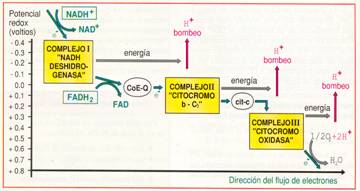
Entre las moléculas de la cadena respiratoria hay transportadores de electrones y transportadores de hidrógeno. Cuando un transportador de hidrógeno es oxidado por un transportador de electrones, los protones quedan libres en la matriz. La disposición ordenada y fija de los transportadores en la membrana permite utilizar la energía liberada en el transporte de electrones para bombear estos protones desde la matriz hacia el espacio intermembrana.
Hay tres puntos en la cadena respiratoria en los que ocurre esta translocación de protones que, debido a la impermeabilidad de la membrana mitocondrial interna, se acumulan en el espacio intermembrana. Así se origina un gradiente electroquímico de protones, es decir, un gradiente de concentración (o lo que es lo mismo, de pH) y un gradiente eléctrico (potencial de membrana), al ser diferente la distribución de H+ a ambos lados de la membrana.
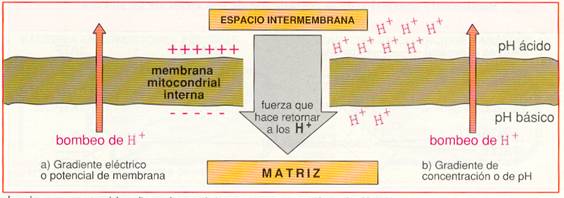
Los dos componentes del gradiente electroquímico se suman para impulsar a los H+ de vuelta a la matriz.
Debido a la impermeabilidad de la membrana interna, el retorno de protones a la matriz sólo puede hacerse a través de la ATP-sintetasa. Esta proteína utiliza la energía acumulada en el gradiente de H+ para fosforilar el ADP y transformarlo en ATP .La hipótesis que explica el acoplamiento de estos dos procesos, uno químico, de oxidación-reducción en la cadena respiratoria, y uno osmótico, de transporte de protones, se conoce con el nombre de hipótesis quimiosmótica y su resultado es la fosforilación oxidativa.
|
|
|
Transporte de electrones, translocación de protones y fosforilación a partir del NADH. |
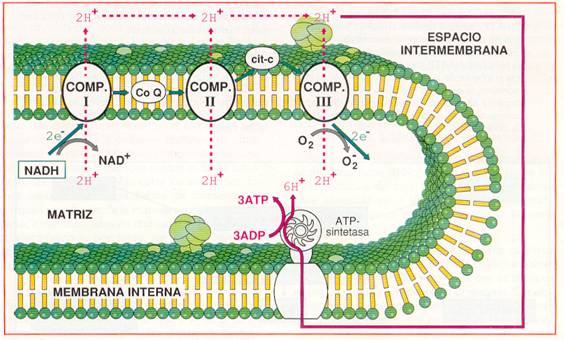
Cada NADH que llega a la cadena respiratoria cede una pareja de electrones, que en su transporte liberan energía suficiente para bombear 6 H+ desde la matriz al espacio intermembrana. Si los electrones proceden del FADH2 sólo se bombean 4 H+.
Por cada 2 H+ que vuelven a la matriz a través de la ATP-sintetasa, se fosforila un ADP. Por tanto, en la cadena respiratoria, se pueden obtener 3 ATP por cada NADH + H+ obtenido en el catabolismo y 2 ATP por cada FADH2.
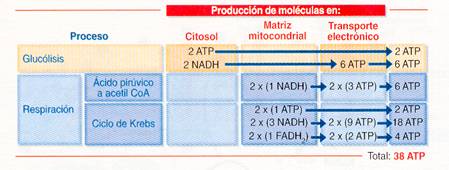
2.5.5.4.2.4 Balance energético del catabolismo de la glucosa.
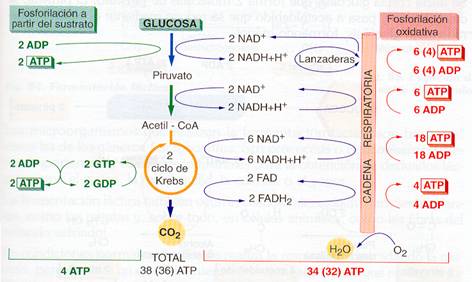
Si admitimos que a partir del NADH + H+ se producen 3 ATP y del FADH2 2 ATP, el balance de la oxidación completa de una molécula de glucosa es el siguiente:
- En la glucólisis se forman 2 ATP, 2 NADH + H+ y 2 piruvato. Dependiendo del tipo de lanzadera que impulse las moléculas de NADH + H+ hacia la cadena respiratoria, éstas rendirán 6 ó 4 moléculas de ATP.
-En la transformación de 2 moléculas de piruvato en 2 de acetil-CoA se liberan 2 NADH que rendirán 6 ATP.
-En el ciclo de Krebs, 2 moléculas de acetil-CoA se oxidan produciendo 2 ATP, 2 FADH2 y 6 NADH. En total rendirán 24 ATP.
Por tanto, en la oxidación completa de la glucosa a CO2 y H2O se produce un máximo de 38 ATP (34 por fosforilación oxidativa y tan sólo 4 por fosforilación a partir del sustrato).
C6H1206 + 6 O2 —> 6 C02 + 6 H2O + 38 ATP
Los 38 ATP por molécula de glucosa suponen un rendimiento máximo que no se suele alcanzar. Si interviene la lanzadera de glicerol-fosfato serían 36 ATP (esta lanzadera el NADH + H+ citosólico pasa a FADH2 mitocondrial); pero, sobre todo hay que tener en cuenta que el gradiente electroquímico de protones de la mitocondria puede usarse para otros fines diferentes de la síntesis de ATP.
|
|
|
Balance energético del catabolismo de la glucosa. |
En el siguiente cuadro también se puede ver resumido:
2.5.5.5. Características generales del anabolismo celular: divergencia metabólica y necesidades energéticas.
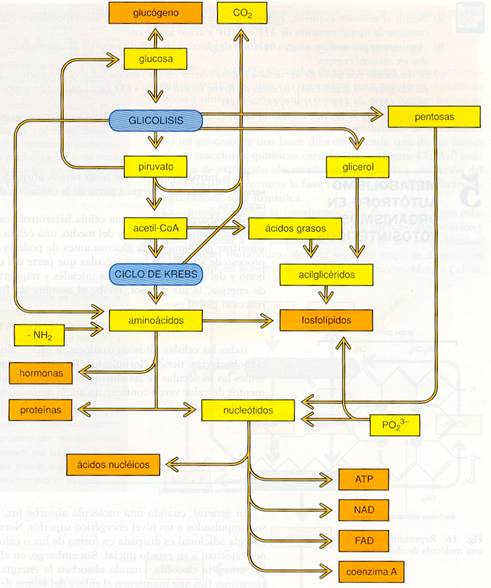
Las reacciones anabólicas son reacciones de síntesis, en general de carácter reductor , que precisan energía. A partir de unos pocos sustratos se pueden formar muchos productos diferentes, por lo que hay divergencia en los productos (Divergencia metabólica).
El anabolismo es la vía constructiva del metabolismo, es decir, la ruta de síntesis de moléculas complejas a partir de moléculas sencillas. Si las moléculas iniciales son inorgánicas (CO2, H2O, etc.) se denomina anabolismo autótrofo, mientras que si son orgánicas (glucosa, aminoácidos, nucleótidos, etc.) Se denomina anabolismo heterótrofo.
- El anabolismo autótrofo se puede realizar mediante la fotosíntesis o mediante la quimiosíntesis.
- El anabolismo heterótrofo se da en todos los organismos, y se realiza de forma similar en todos ellos. Su objeto es la síntesis de reservas energéticas y crear estructuras para que el organismo pueda crecer o simplemente, para que pueda renovar sus estructuras deterioradas.
Vamos a ver algunos principios básicos del anabolismo o biosíntesis celular:
1º Cada célula,en general, sintetiza sus propias proteínas, ácidos nucleicos, lípidos, polisacáridos y otras moléculas complejas, de modo que no las recibe de otras células.
2º Cada uno de los pasos de la biosíntesis es catalizado por una enzima particular.
ass=MsoNormal style='text-align:justify;mso-layout-grid-align:none; text-autospace:none'>
3º Aunque algunos pasos de la secuencia biosintética se realizan sin el uso de fosfatos ricos en energía, la síntesis global de esas moléculas demanda el consumo de energía (necesidades energéticas)
4º Para los procesos biosintéticos se utilizan relativamente pocas sustancias como materia, prima entre las cuales se encuentran acetil-Co A, ribosa, piruvato y glicerol (divergencia metabólica).
5º Estos procesos anabólicos, no son, en general, la simple inversión de procesos de degradación de las moléculas, sino que comprenden uno o más pasos que difieren de las etapas del proceso catabólico. Estos pasos están bajo el control de enzimas diferentes, lo que permite que sean mecanismos diferentes de control los que gobiernen la síntesis y degradación de la misma molécula compleja.
6º Todas las moléculas constituyentes de las células se encuentran en un estado dinámico en el que son degradadas y sintetizadas constantemente.
7º Los procesos biosintéticos no sólo comprenden la formación de los componentes macromoleculares a partir de precursores simples, sino también su ensamblaje para formar los varios tipos de membranas que componen los limites externos de la célula y los orgánulos intracelulares.
8º La célula esté o no en crecimiento utiliza una fracción considerable de energía en el proceso biosintético.
9º Muchos de los pasos de los procesos de biosíntesis consisten en la formación de enlaces peptídicos, glucosídicos y estéricos. Aunque esos enlaces se rompen por hidrólisis, no se forman a través de reacciones en las que se pierde agua.
|
|
|
Esquema general del anabolismo en células heterótrofas. |
2.5.5.5.1. Concepto e importancia biológica de la fotosíntesis.
La fotosíntesis es un complejo proceso anabólico, mediante el cual los seres vivos poseedores de clorofila y otros pigmentos (algunas bacterias, algas y plantas), incorporan materia (CO2, H2O) y energía (solar), transformando estos compuestos minerales en otros compuestos orgánicos (glúcidos, etc.).
El proceso tiene lugar en los cloroplastos, gracias a la presencia de una serie de pigmentos capaces de aumentar su nivel energético al incidir sobre ellos la energía luminosa. Estos pigmentos son las clorofilas, carotenoides y ficobilinas.
La ecuación global de este proceso es la siguiente:
Clorofila
6 CO2 + 12 H2O ————> C6H12O6 + 6 H2O + 6 O2
Luz
De múltiples observaciones experimentales, se deduce que para que se realice la fotosíntesis se requiere:
- presencia de la luz;
- concurso de la clorofila;
-dióxido de carbono (CO2);
- agua.
Como consecuencia del fenómeno hay:
- desprendimiento de O2;
- formación de compuestos orgánicos (síntesis).
Asimismo se ha visto que los productos iniciales son: CO2 y H2O los productos finales compuestos orgánicos y O2.
Importancia biológica de la fotosíntesis.
La fotosíntesis es seguramente el proceso bioquímico más importante de la Biosfera por varios motivos:
a) La síntesis de materia orgánica a partir de la inorgánica se realiza fundamentalmente mediante la fotosíntesis; luego irá pasando de unos seres vivos a otros mediante las cadenas tróficas, para ser transformada en materia propia por los diferentes seres vivos.
b) Produce la transformación de la energía luminosa en energía química, necesaria y utilizada por todos los seres vivos.
c) Enla fotosíntesis se libera oxígeno como subproducto, que es necesario para ser utilizado en la respiración aerobia como oxidante.
d) La fotosíntesis fue causante del cambio producido en la atmósfera terrestre primitiva, que era anaerobia y reductora.
e) De la fotosíntesis depende también la energía almacenada en combustibles fósiles como carbón, petróleo y gas natural.
f) Elequilibrio necesario entre seres autótrofos y heterótrofos no sería posible sin la fotosíntesis.
Se puede concluir que la diversidad de vida existente en la Tierra depende principalmente de la fotosíntesis.
2.5.5.5.2. Etapas de la fotosíntesis y su localización.
La fotosíntesis comprende numerosas reacciones químicas. De ellas, unas utilizan necesariamente la energía lumínica; otras no. Basándose en esto, para estudiar la fotosíntesis se consideran dos fases:
- La fase lumínica: conjunto de reacciones que requieren luz.
- La fase oscura: conjunto de reacciones que no requieren luz.
Estas dos fases se realizan simultáneamente.
FOTOFOSFORILACION y SINTESIS DE PODER REDUCTOR (fase luminosa).
Las reacciones dependientes de la luz o luminosas de la fotosíntesis empiezan con la clorofila. Las moléculas de clorofila y sus aceptores de electrones asociados (pigmentos fotosintetizadores) se localizan en unidades llamadas fotosistemas.Cada fotosistema es un conjunto molecular muy complejo que está formado por una antena colectora y un centro de reacción. La antena colectora es un grupo de varios cientos de pigmentos (clorofilas, carotenoides, etc.) que está unido a la membrana tilacoidal a través de unas proteínas especiales. Su función es dirigir la energía luminosa recogida hacia una clorofila especial llamada clorofila del centro de reacción.
Existen dos tipos de fotosistemas (fotosistema.I y fotosistema II)que se diferencian por su distinta composición y por captar energía luminosa en dos longitudes de onda (l) diferentes.
La absorción de la luz se efectúa de forma fotónica, y produce en el pigmento un cambio de su distribución electrónica; el pigmento está excitado.
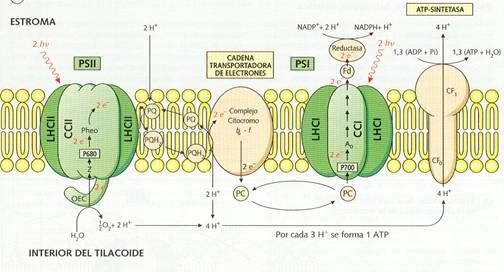
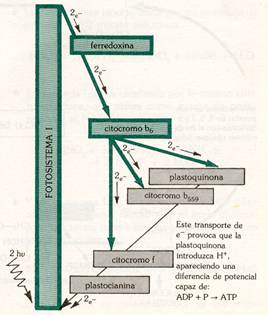
Al incidir la luz (2 fotones) sobre los fotosistemas I y II, sufren la perdida de dos electrones cada uno. Los electrones perdidos por el fotosistema I son captados por un transportador electrónico (ferredoxina) que se reduce; éste transportador, al volverse a oxidar, reduce al NADP a NADPH2 este último es el reductor necesario para la reducción del CO2 en la fase oscura. El fotosistema I ha de recuperar los electrones perdidos y lo hace al incorporarse los electrones del fotosistema II; éstos al saltar del fotosistema II, son tomados por el citocromo b, a través de la plastoquinona que se reduce en primer lugar y luego se oxida. Al oxidarse el citocrorno b cede los electrones al citocromo f; en este paso se producen dos moléculas de ATP (energía química potencial), a partir de ADP + Pi; .El citocromo f vuelve a oxidarse, dando sus electrones al fotosistema I a través de la plastocianina.
El problema que se plantea es de cómo se recuperan los electrones del fotosistema II, ya que si este se sigue excitando, llegaría a desaparecer. Estos electrones provienen de la ruptura de la molécula de agua por la luz, con desprendimiento de O2; esta reacción de denomina fotolisis del agua.
Esquema de la fase lumínica de la fotosíntesis.
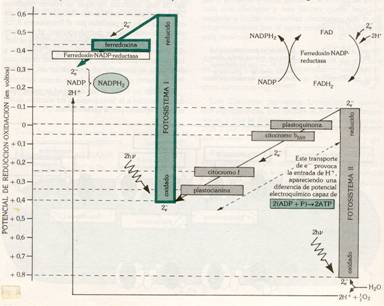
En este proceso fotoquímico se pueden diferenciar por tanto, claramente tres partes acopladas entre sí.
En primer lugar, la ruptura de la molécula de agua por la luz con desprendimiento de O2 y electrones, que pasarán a restablecer el equilibrio del fotosistema II; es la fotolisis del agua:
H2O ——————> ½ O2 + 2H+ + 2e-
Tiene lugar en el interior de los tilacoides, en la parte interna de su membrana.
En segundo lugar, la formación de ATP por fosforilación de ADP con Pi en presencia de iones Mg++, gracias a la energía fotónica; es la fotofosforilación acíclica:
n ADP + n Pi ¾¾¾ > n ATP
Se produce en la parte externa de la membrana del tilacoide.
Y, en tercer lugar, la formación del NADPH2 (poder reductor) por reducción del NADP, al tomar los electrones perdidos en el fotosistema I; es la fotorreducción del NADP ó síntesis de poder reductor:
NADP +2H+ +2e- ¾¾¾ > NADPH2
Esta parte del proceso también se produce en contacto con la parte externa de la membrana del tilacoide.
Membrana de los tilacoides.
Las bacterias fotosintetizantes realizan la fase fotoquímica exclusivamente en el fotosistema I;el cual cede dos electrones a la ferredoxina y esta directamente los cede al citocromo b6al oxidarse ella. Este citocromo puede cederlos a la plastoquinona, al citocromo b3 o al citocromo f, que devuelven los electrones al fotosistema I a través de la plastocianina. De esta manera se forma un ciclo en que no ocurre la reducción del NADP, ni la fotolisis del agua; únicamente se produce la fotofosforilación, que en este caso se denomina cíclica, de manera que este mecanismo só1o sirve para formar ATP a partir de ADP, Mg++, Pi,y energía captada por el fotosistema I.
|
|
|
Fotofosforilidación cíclica. |
Al carecer las bacterias de cloroplastos, la fase fotoquímica tiene lugar en las membranas internas de los mesosomas.
ASIMILACION DEL CO2: CICLO DECALVIN (fase oscura).
La asimilación del CO2, no precisa de la luz, produciéndose por tanto en la llamada fase oscura de la fotosíntesis. Esta fase oscura consiste en un conjunto de reacciones en las que se reduce el CO2, incorporándose a un azúcar simultáneamente para formar otro monosacárido gracias al ATP y al NADPH2 formados en la fase luminosa. La fase oscura ocurre en el estroma de los cloroplastos de las plantas superiores o en el protoplasma de bacterias y cianofíceas.
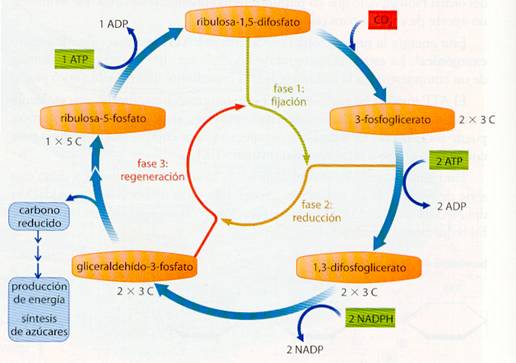
La fijación del CO2 a nivel de azúcar (o de otro compuesto) puede considerarse que ocurre en tres fases distintas, donde las tres están formando un círculo, por lo que también se conoce con el nombre de Ciclo de Calvin, su descubridor.
Estas fases son:
- Carboxilación o fijación. Mediante la enzima ribulosa difosfato carboxilasa, un compuesto con 5 carbonos, la ribulosa-l, 5-difosfato, se combina con el CO2, formándose un compuesto muy inestable de 6 átomos de carbono que se rompe inmediatamente en dos moléculas de 3-fosfoglicerato.
- Reducción. El 3-fosfoglicerato es, en primer lugar, fosforilado con el correspondiente consumo energético de ATP, formándose 1,3-difosfoglicerato. Seguidamente, gracias al NADPH, se reduce a gliceraldehído-3-fosfato.
- Recuperación o regeneración. De cada 6 moléculas de gliceraldehído-3-fosfato que se forman, una se considera el rendimiento neto del ciclo. Las otras 5 sufren una serie de transformaciones consecutivas, con diferentes monosacáridos fosfatados, en las que también se consume ATP, para regenerar la ribulosa-l,5-difosfato, con la que se cierra el ciclo.
|
|
|
Ciclo de Calvin |
El problema de la fotorrespiración
La enzima ribulosa difosfato carboxilasa puede funcionar en dos sentidos diferentes. En el ciclo de Calvin cataliza la carboxilación de la ribulosa-1,5-difosfato, pero también puede provocar la oxigenación de esta molécula.
El que actúe como carboxilasa u oxigenasa depende de las concentraciones relativas CO2 y del O2 en el interior de la célula.
La oxigenación de la ribulosa-l,5-difosfato recibe el nombre de fotorrespiración, ya que depende de la luz, pero se parece a la respiración pues consume O2 y libera CO2. Tiene lugar en tres compartimentos celulares: cloroplastos, peroxisomas y mitocondrias.
|
|
|
Esquema de la fotorrespiración |
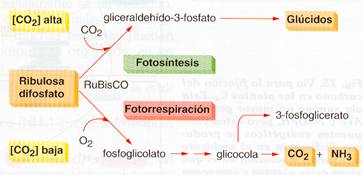
El resultado de la fotorrespiración es que únicamente se consume materia orgánica, sin formación de ATP, y parece claramente perjudicial para planta.
|
|
|
Esquema general de la fotosíntesis. |
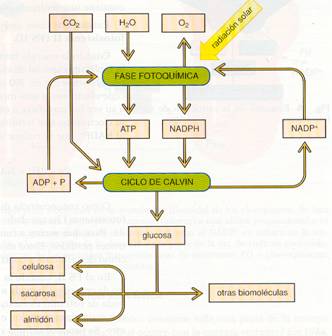
Balance de la fotosíntesis.
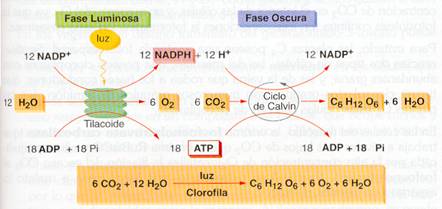
En la fase luminosa se produce el ATP y el NADPH necesarios para, en la fase oscura, reducir el CO2 a materia orgánica. Si, por ejemplo, se considera la síntesis de una molécula de glucosa (C6H12O6), se observa que son necesarios 6 CO2 y 12 H20. Esta agua libera sus 6 O2 a la atmósfera, durante la fase luminosa, y aporta los 12 hidrógenos de la glucosa y los 12 hidrógenos necesarios para pasar los 6 O2 sobrantes del CO2 a H2O. Como intervienen 24 hidrógenos, aparecen 24 H+ y 24 e-, y como cada electrón precisa el impacto de dos fotones, uno en el PSI y otro en el PSII, se necesitan 48 fotones (hv). En el ciclo de Calvin se precisan, por cada CO2 incorporado, 2 NADPH y 3 ATP; así pues, para una glucosa son necesarios 12 NADPH y 18 ATP.
|
|
|
Balance global de la fotosíntesis |
Fijación fotosintética del nitrógeno y del azufre
a) Síntesis de compuestos orgánicos nitrogenados. La reducción de los iones nitrato que se encuentran disueltos en el suelo se realiza, gracias al ATP y al NADPH obtenidos en la fase luminosa, en tres etapas:
1. Los iones nitrato (NO3-) son reducidos a iones nitrito (NO2-) por la enzima nitrato reductasa, gastando un NADPH.
2. Los nitritos son reducidos a amoniaco (NH3) por la enzima nitrito reductasa, gastando un NADPH.
3. A continuación, el amoniaco obtenido, que resulta tóxico para la planta, es rápidamente captado por el ácido α-cetoglutárico, dando lugar al ácido glutámico. De este modo pasa a formar parte de la materia orgánica de la célula. Esta reacción es catalizada por la enzima glutamato sintetasa y precisa el gasto de un ATP. A partir del ácido glutámico, los átomos de nitrógeno pueden pasar, en forma de grupo amino, a otros cetoácidos y dar lugar a otros aminoácidos. Algunas bacterias y cianobacterias pueden fijar el nitrógeno atmosférico, N2.
b) Síntesis de compuestos orgánicos con azufre. A partir del NADPH y del ATP de la fase luminosa, se reduce el ión sulfato (SO42-) a ión sulfito (SO32-) y luego a sulfuro de hidrógeno (H2S). Éste, al combinarse con la acetilserina, da lugar al aminoácido cisteína, con lo que pasa a formar parte de la materia orgánica celular.
Factores que influyen en la fotosíntesis.
El rendimiento o eficacia de la fotosíntesis se puede medir en función de la concentración de CO2 asimilada por unidad de tiempo o con relación a la cantidad de O2 desprendido. Los principales factores de carácter ambiental que influyen en velocidad de la fotosíntesis son:
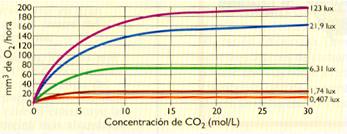
- Concentración de CO2. Siempre que se mantenga constante y suficientemente elevada la intensidad luminosa, el aumento de CO2 en el aire incrementa el rendimiento de la fotosíntesis. Este aumento se produce hasta alcanzar un valor de asimilación máxima, específico para cada organismo, por encima del cual el rendimiento se estabiliza.
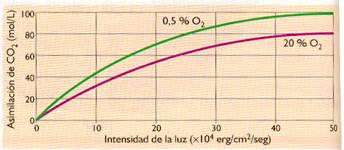
- Concentración de O2. Cuando aumenta la concentración de O2 disminuye el rendimiento fotosintético debido a la competencia que se establece entre los sustratos de la enzima rubisco, favoreciéndose la fotorrespiración.
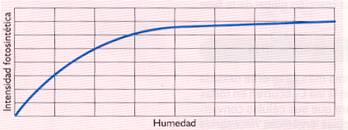
- Humedad. La concentración de agua tanto en el suelo como en el ambiente influye de manera determinante en el rendimiento de la fotosíntesis, debido a que se trata de una materia prima. Al disminuir el grado de humedad se cierran los estomas para evitar pérdidas de agua, reduciéndose la asimilación de CO2.
- Temperatura. Para intensidades luminosas altas, el rendimiento de la fotosíntesis aumenta con la temperatura hasta alcanzar un valor máximo, siendo este el de la temperatura óptima para la actuación de las enzimas. Los valores más alejados provocan la desnaturalización de las enzimas y el rendimiento fotosintético disminuye considerablemente.
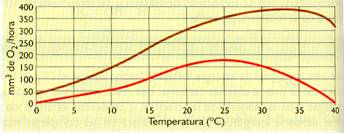
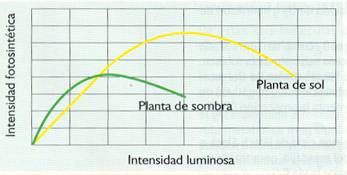
- Intensidad luminosa. Para intensidades luminosas relativamente bajas, la fotosíntesis es proporcional a la intensidad de luz; pero si esta va aumentando, llega un momento en el que el rendimiento se estabiliza según las características óptimas de los pigmentos de cada especie.
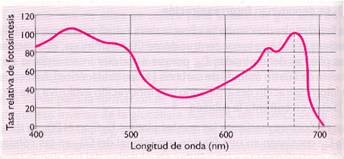
- Tipo de luz. El rendimiento óptimo de la fotosíntesis se realiza con luz roja o azul. Con luz roja de 680 nm la absorción de la luz va acompañada de absorción de gran cantidad de energía. Al disminuir la longitud de onda se produce absorción de más cantidad de energía. Cuando la longitud de onda es superior a 680 nm no actúa el fotosistema II, con lo que el rendimiento disminuirá por no poder realizarse la fase oscura de la fotosíntesis.
2.5.5.5.3. Fotosíntesis y evolución.
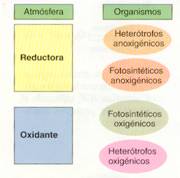
La atmósfera primitiva era reductora y, para los primeros organismos, el oxígeno era una sustancia tóxica que podía sustraer electrones de los componentes moleculares de sus células. Los seres vivos obtenían la energía degradando la glucosa a ácido pirúvico (glucólisis anaeróbica): eran heterótrofos anoxigénicos. Pero los compuestos orgánicos se agotaron en seguida, ya que eran generados por procesos geoquímicos muy lentos.
Algunos organismos desarrollaron la capacidad de reducir fuentes de carbono muy abundantes, como el CO2, utilizando la energía luminosa (fotosíntesis). Esto supuso una auténtica revolución.
La vida en la Tierra depende de la energía solar y su conversión en energía química requiere una transferencia de electrones entre moléculas. Así, las células necesitan una fuente de electrones para vivir.
Las actuales bacterias verdes del azufre utilizan la luz para transferir H+ y electrones desde el H2S hasta el NADPH, creando el poder reductor necesario para la fijación del carbono. Antepasados de estas bacterias pudieron ser los primeros organismos fotosintéticos, pero su distribución debía de ser muy limitada. Las bacterias anoxigénicas fotosintéticas actuales sólo existen en fuentes sulfúreas, en fondos lacustres y en ambientes similares.
Otra auténtica revolución para los organismos fotosintéticos fue la aparición de células capaces de utilizar el agua como dador electrónico, lo cual permitió invadir ambientes diversos. Hace aproximadamente 3.000 millones de años, algunas células fotosintéticas (cianobacterias) desarrollaron la facultad de disociar pares de moléculas de agua en electrones, protones y oxígeno. Eran organismos fotosintéticos oxigénicos. Los electrones y protones resultaban energéticamente útiles, y el oxígeno era un producto de desecho. El incremento de oxígeno fue tal, que la primitiva atmósfera reductora se sustituyó progresivamente por otra oxidante.
La fotosíntesis proporcionaba compuestos orgánicos que podían ser utilizados por otros organismos (heterótrofos), y oxígeno. El uso del oxígeno en la degradación de compuestos orgánicos aumentó enormemente el rendimiento energético de este proceso (respiración celular) y permitió la rápida evolución de grandes grupos de organismos aerobios pluricelulares, tanto autótrofos como heterótrofos. En la actualidad, la fotosíntesis es la fuente de oxígeno atmosférico, el cual es esencial para la mayoría de losseres vivos.
|
|
|
Grupos de organismos desarrollados según el tipo de atmósfera existente. |
2.5.5.5.4. Quimiosíntesis.
La quimiosíntesis es un proceso metabólico realizado únicamente por algunas bacterias autótrofas. Consiste en la obtención de energía (ATP) a partir de la oxidación de diversas sustancias inorgánicas; y el posterior uso de esa energía para transformar sustancias inorgánicas en compuestos orgánicos.
Así, en la quimiosíntesis se diferencian dos fases:
- En la primera fase, la oxidación de sustancias inorgánicas como NH3, NO2-, H2, H2S, S, Fe2+, etc., permite generar un gradiente de protones, entre el citoplasma y el espacio periplasmático, capaz de impulsar la ATP sintasa, formándose ATP mediante un proceso de fosforilación oxidativa. Parte de ese ATP hará posible un flujo inverso de electrones, gracias al que se obtiene NADH (raramente NADPH).
- La segunda fase es coincidente o muy semejante a la fase oscura de la fotosíntesis. Así, la fijación del CO2 ocurre generalmente a través del ciclo de Calvin; y el nitrógeno se obtiene a partir de la reducción de nitratos
|
|
|
Etapas de la quimiosíntesis. |
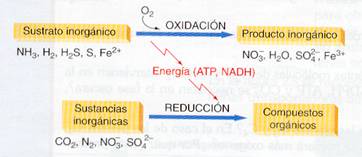
Clases de bacterias quimiosintéticas
Lo peculiar de la quimiosíntesis es la primera fase, la obtención de energía mediante la oxidación de sustancias inorgánicas. Por ello, las bacterias quimiosintéticas se clasifican atendiendo al sustrato inorgánico que utilicen.
· Bacterias del nitrógeno. Son bacterias comunes en los suelos e imprescindibles para el correcto desarrollo del ciclo del nitrógeno. Se diferencian dos grupos:
- Bacterias que transforman el amoníaco en nitritos. Por ejemplo Nitrosomonas.
2 NH3 + 3O2 ——> 2 NO2- + 2 H+ + 2 H2O + energía
- Bacterias que transforman los nitritos en nitratos. Por ejemplo Nitrobacter.
2 NO2- + 1/2 O2 ——> 2 NO3- + energía
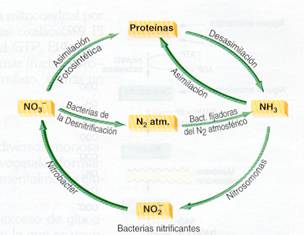
Ambos grupos de bacterias trabajan ecológicamente unidos, uno a continuación del otro, y hacen posible la nitrificación del amoníaco.
|
|
|
Ciclo ecológico del nitrógeno |
· Bacterias incoloras del azufre. Viven en las aguas residuales de las poblaciones, en emanaciones hidrotermales y en otros ambientes con H2S y otros derivados del azufre, oxidando estos sustratos para obtener energía. No se deben confundir con las sulfobacterias verdes o purpúreas, que utilizan H2S, pero son fotosintéticas.
H2S + ½ O2 ——> S + H2O + energía
2 S + 2 H2O + 3 O2 ——> 2 S042- + 4 H+ + energía
· Bacterias del hierro. Viven en aguas ricas en sales ferrosas que oxidan a férricas. Por ejemplo Ferrobacillus.
4 Fe2+ + 4 H+ + O2 ——— > 4 Fe3+ + 2 H2O + energía
· Bacterias del hidrógeno y del metano. Utilizan estos sustratos como fuente reductora.
H2 + ½ O2 ——> H2O + energía
CH4 + 2 O2 ——> CO2 + 2 H2O + energía
Las bacterias del metano, junto con algunas bacterias del azufre, abundan en los fondos marinos, donde se producen emanaciones volcánicas, constituyendo la base alimenticia de unos ecosistemas muy peculiares, situados a más de 2.000 m de profundidad.
2.5.5.6. Integración del catabolismo y del anabolismo
El conjunto del metabolismo depende de dos procesos antagónicos en sus términos extremos: la fotosíntesis que tiene lugar en los cloroplastos y la respiración que termina en las mitocondrias. En el conjunto de la Biosfera estos dos procesos deben permanecer en equilibrio y regulan dos ciclos biogeoquímicos fundamentales: el del oxígeno y el del carbono.
En las células eucariotas, las mitocondrias son orgánulos clave en los que no sólo terminan casi todas las rutas catabólicas y tiene lugar la fosforilación oxidativa sino que también comienza en ellas el anabolismo de la glucosa y de los lípidos. La transformación del piruvato en acetil coA es irreversible, lo que determina que en las células animales los ácidos grasos no se puedan convertir en glúcidos.