2.3. Célula eucariota. Componentes estructurales y funcionales.
2.3. Célula eucariota. Componentes estructurales y funcionales.
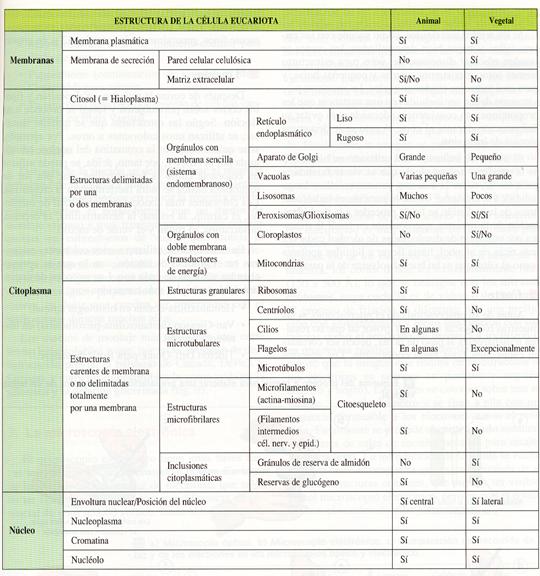
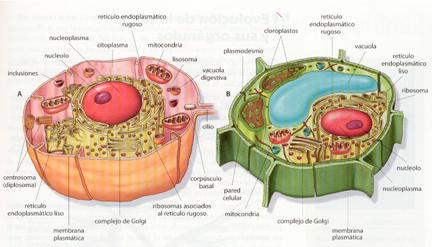
Las células eucariotas, pese a tener unas estructuras comunes a todas ellas, presentan dos tipos de organización general según se encuentren constituyendo organismos animales o vegetales. Se puede, pues, hablar de células vegetales y de células animales. Ambos tipos poseen membrana plasmática, citoplasma con sistema endomembranoso, mitocondrias, lisosomas, peroxisomas, vacuolas, citoesqueleto y núcleo con envoltura nuclear. Sin embargo, las diferencias acumuladas a lo largo de millones de años de evolución independiente han causado que las células de los vegetales posean estructuras que faltan en las animales, y viceversa. A continuación se citan algunas de estas diferencias:
a) En las células de los vegetales destaca la presencia de una pared de secreción gruesa de celulosa, la existencia en general de una vacuola grande que desplaza el núcleo desde el centro a un lado, la presencia de plastos que almacenan el polisacárido almidón y que, si son estimulados por la luz, se enriquecen en clorofila y se convierten en cloroplastos fotosintéticos.
b) En las células de los animales, si hay membrana de secreción, es de mucopolisacáridos, la denominada matriz extracelular, las vacuolas son pequeñas, el núcleo suele estar en el centro, hay un diplosoma formado por dos centríolos, puede presentar cilios, o flagelos o emitir pseudópodos, y el polisacárido con función de reserva energética no es el almidón sino el glucógeno.
Diferencias entre células de los animales y de los vegetales.
2.3.1. Membranas celulares: composición, estructura y funciones.
La membrana plasmática es una estructura que rodea y limita completamente a la célula y constituye una «barrera» selectiva que controla el intercambio de sustancias desde el interior celular hacia el medio exterior circundante, y viceversa.
La membrana plasmática posee la misma estructura en todas las células. En cortes ultrafinos aparece como dos bandas oscuras separadas por una banda clara, con un espesor de 7,5 nm. Esta organización es común, además, al resto de las membranas biológicas constituyentes o limitantes de los orgánulos celulares, por lo que se denomina unidad de membrana (o membrana unitaria).
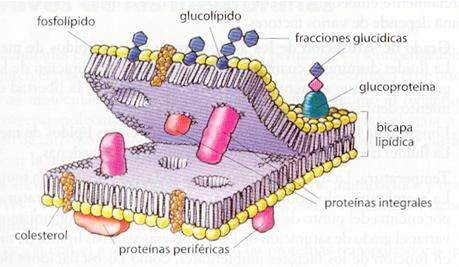
La estructura trilaminar observada en la unidad de membrana se corresponde con una bicapa lipídica con proteínas embebidas. Los lípidos se disponen en una bicapa con las zonas hidrófilas (grupos polares) hacia fuera, mientras que las zonas hidrófobas quedan enfrentadas hacia el interior. Las membranas presentan, por tanto, dos caras: una cara externa y una cara interna que, en el caso de la membrana plasmática, está en contacto con el citoplasma celular. Las proteínas pueden estar asociadas a la cara interna o externa, o ser transmembranales (atraviesan la membrana totalmente).
ESTRUCTURA DE LA MEMBRANA. MODELO DEL MOSAICO FLUIDO
Con los datos ofrecidos por la microscopía electrónica y los análisis bioquímicos se han ido elaborando varios modelos a lo largo del desarrollo de la biología celular. En la actualidad, el modelo más aceptado es el propuesto por Singer y Nicholson (1972), denominado modelo del mosaico fluido, que presenta las siguientes características:
· Considera que la membrana es como un mosaico fluido en el que la bicapa lipídica es la red cementante y las proteínas están embebidas en ella, interaccionando unas con otras y con los lípidos. Tanto las proteínas como los lípidos pueden desplazarse lateralmente.
· Los lípidos y las proteínas integrales se hallan dispuestos en mosaico.
· Las membranas son estructuras asimétricas en cuanto a la distribución de todos sus componentes químicos: lípidos, proteínas y glúcidos
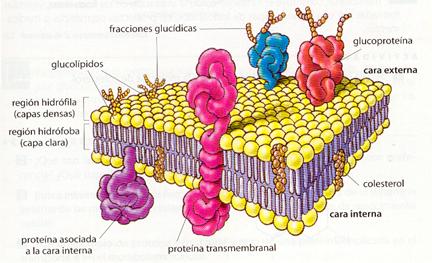
Estructura de la membrana plasmática según el modelo del mosaico fluido.
COMPOSICIÓN.
La membrana está compuesta fundamentalmente por lípidos y proteínas, y en menor cantidad por glúcidos. Su composición relativa se determinó por primera vez en eritrocitos de rata ( 40% de lípidos y 60% de proteínas ). Posteriormente se comprobado que dicha proporción es muy similar en el resto de las células aunque puede variar en función del tipo celular; por ejemplo, en los hepatocitos de rata la proporción es de un 58% de lípidos y un 42% de proteínas, mientras que en las fibras nerviosas las proteínas alcanzan menos del 25%, y en músculo esquelético de rata, el 65% del total.
Lípidos de membrana
Los lípidos de membrana pertenecen fundamentalmente a tres categorías: fosfolípidos, glucolípidos y esteroles.
- Fosfolípidos. Son los lípidos más abundantes en las membranas biológicas. Presentan una zona hidrófila, que constituye las denominadas cabezas polares (glicerina o glicerol en los fosfoglicéridos), y una zona hidrófoba (ácidos grasos), que forma la colaapolar. Los fosfo1ípidos poseen, por tanto, un carácter anfipático.
- Glucolípidos. Son muy semejantes a los fosfolípidos, pero contienen oligosacáridos. En las células animales suelen ser derivados de esfingolípidos. En las células vegetales y procariotas, sin embargo, los glucolípidos derivan de los fosfog1icéridos. Sólo aparecen en la cara externa de la membrana plasmática.
- Esteroles. Derivados del colesterol y presentes en la membrana plasmática de las células eucariotas, son más abundantes, por lo general, en las células animales.
La membrana plasmática no es una estructura estática: sus componentes tienen posibilidad de movimiento, lo que le proporciona una cierta fluidez.
Los movimientos que pueden realizar los lípidos son:
· De rotación: supone el giro de la molécula lipídica en torno a su eje mayor. Es muy frecuente y el responsable, en gran medida, de los otros dos movimientos.
· De difusión lateral: las moléculas lipídicas pueden difundirse libremente de manera lateral dentro de la bicapa. Es el movimiento más frecuente.
· Flip-flop: es el movimiento de la molécula lipídica de una monocapa a la otra gracias a unas enzimas llamadas lipasas. Es el movimiento menos frecuente, por ser muy desfavorable energéticamente.
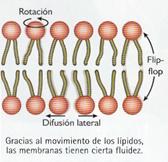
La fluidez o viscosidad es una de las características más importantes de las membranas. Depende de factores como la temperatura (la fluidez aumenta al incrementarse la temperatura), la naturaleza de los lípidos (la presencia de lípidos insaturados y de cadena corta favorece el aumento de la fluidez) y la presencia de colesterol (endurece las membranas, reduciendo su fluidez y permeabilidad). De la fluidez dependen importantes funciones de la membrana, como el transporte, la adhesión celular o la función inmunitaria. Por ello, las membranas poseen mecanismos de adaptación homeoviscosa encargados de mantener la fluidez.
Proteínas de membrana
Las proteínas asociadas a la membrana pueden cumplir un papel meramente estructural, funciones de reconocimiento y adhesión, o bien estar implicadas en el transporte y el metabolismo celular. Según su grado de asociación a la membrana se clasifican en dos grupos: integrales y periféricas.
· Integrales. Estas proteínas se asocian a la membrana mediante enlaces hidrófobos. Sólo pueden separarse de la membrana si se destruye la bicapa (por ejemplo, con detergentes neutros), Dentro de este grupo existen proteínas transmembranales y proteínas asociadas a la cara externa o a la cara interna de la membrana. Algunas proteínas presentan hidratos de carbono unidos a ellas covalentemente (glucoproteínas) y se disponen siempre en el lado externo de la membrana, como los glucolípidos.
· Periféricas. Son proteínas unidas a la membrana por enlaces de tipo iónico y se separan de ella con facilidad (por ejemplo! con soluciones salinas, que mantienen intacta la bicapa). Aparecen principalmente en la cara interna de la membrana. En este grupo no existen proteínas transmembranales.
Como se ha visto, la composición de los lípidos y de las proteínas es diferente en las dos caras de la membrana. Por esta razón se dice que las membranas son asimétricas , es decir, se pueden diferenciar las caras interna y externa en función de su composición.
Asimetría de la membrana plasmática.
Exteriormente a la membrana algunas células presentan un glicocáliz , compuesto por glucoproteínas, que pueden interaccionar o estar parcialmente incluidas en la membrana plasmática, y por glucolípidos. Esta matriz extracelular es importante en los procesos de reconocimiento e interacción entre las células y los tejidos.
FUNCIONES DE LA MEMBRANA CELULAR.
La función principal de la membrana plasmática consiste en limitar la célula y, por tanto, en separar el citoplasma y sus orgánulos del medio que los rodea. Este papel no es pasivo, ya que la membrana actúa como una barrera selectiva para el intercambio y el transporte de sustancias. La membrana celular cumple, además, otras funciones esenciales:
- Producción y control de gradientes electroquímicos, ya que en ella se localizan cadenas de transporte y proteínas relacionadas con los mismos.
- Intercambio de señales entre el medio externo y el medio celular.
- División celular: la membrana está implicada en el control y desarrollo de la división celular o citocinesis.
- Inmunidad celular: en la membrana se localizan algunas moléculas con propiedades antigénicas, relacionadas, por ejemplo, con el rechazo en trasplantes de tejidos u órganos de otros individuos.
- Endocitosis y exocitosis: la membrana está relacionada con la captación de partículas de gran tamaño (endocitosis) y con la secreción de sustancias al exterior (exocitosis).
DIFERENCIACIONES DE LA MEMBRANA.
Van dirigidas al desempeño de una función concreta y consistentes en algún tipo de alteración morfológica del contorno de la célula en cualquiera de sus superficies:
- Superficie apical (que da hacia la luz del conducto): son típicas las microvellosidades de algunas células epiteliales. Se tratan de evaginaciones con forma de dedo de guante que aumentan la superficie de absorción intestinal.
- Superficie basal (lado opuesto a la luz del conducto): también destacan las células epiteliales, concretamente las que en el riñón presentan invaginaciones que aumentan la superficie de reabsorción de agua en el tubo contorneado proximal de las nefronas.
- Superficie lateral: son las denominadas uniones intercelulares que posibilitan las interacciones entre células vecinas. Son de varios tipos: estrechas o impermeables, que no dejan espacio intercelular alguno, comunicantes o en hendidura, que dejan un reducido espacio intercelular, y adherentes o desmosomas, que, aunque con un espacio intercelular mayor, implican una fuerte unión mecánica entre las células.
2.3.2. Pared celular en células vegetales.
Las células vegetales poseen una envuelta externa a la membrana plasmática, altamente organizada y rígida, que constituye la pared celular y cumple las siguientes funciones:
- Confiere rigidez al vegetal y contribuye al mantenimiento de la forma celular.
- Une las células adyacentes, conectando las células de los tejidos vegetales.
- Posibilita el intercambio de fluidos y la comunicación intercelular.
- Permite a las células vegetales vivir en el medio hipotónico de la planta, impidiendo que éstas se hinchen y lleguen a estallar.
- Impermeabiliza la superficie vegetal en algunos tejidos gracias a la cutina y la suberina para evitar las pérdidas de agua.
- Sirve de barrera al paso de agentes patógenos.
Estructura y composición
La pared celular tiene dos componentes principales:
- Un componente cristalino, constituido por fibras de celulosa .
- Un componente amorfo, formado por una matriz de pectinas , hemicelulosa, agua, sales minerales y, en algunos casos, ligninas.
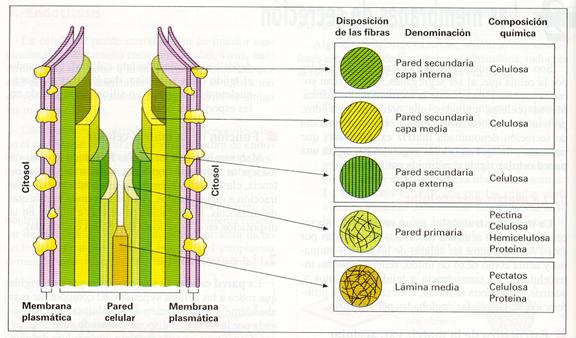
En las células diferenciadas, la pared celular aparece como una estructura gruesa compuesta por varias capas que se van depositando a medida que madura la célula. Estas capas son:
· Lámina media . Es la capa más externa y la primera que se forma después de la división celular, y puede ser compartida por las células adyacentes de un tejido. Está integrada por pectinas y proteínas, que se unen posteriormente a iones Ca2+.
· Pared primaria. Se trata de una gruesa capa de estructura fibrilar, situada por debajo de la lámina media hacia el interior de la célula. Está constituida, fundamentalmente, por largas fibras de celulosa cohesionadas por polisacáridos (hemicelulosa y pectinas) y glucoproteínas.
· Pared secundaria. Es la capa más interna y se encuentra por debajo de la pared primaria en algunos tipos especiales de células vegetales (tejidos de soporte o vasculares). Consta de una o varias capas fibrilares, semejantes en composición a la pared primaria, aunque contienen celulosa en mayor proporción y carecen de pectinas. En estas capas fibrilares, las miofibrillas de celulosa se ordenan paralelamente dando lugar a varias capas con diferente ordenación.
En ocasiones, entran a formar parte de su composición polímeros, como la lignina (xilema y esclerénquima), ceras y cutina (haz de las hojas) o suberina (corcho).
En algunas paredes se observan también inclusiones minerales, principalmente carbonatos y sílice.
Composición de la pared celular.
La pared celular de las células vegetales no es continua, ya que presenta una serie de canales, los plasmodesmos , que permiten la comunicación y e intercambio de líquidos con sustancias disueltas y otras moléculas entre células adyacentes. Estos canales se forman en la división celular, durante la síntesis de la pared, y aparecen en todas las células jóvenes.
Los plasmodesmos pueden encontrarse en ocasiones en depresiones de la pared primaria (menos engrosada), en las que también se inhibe el depósito de pared secundaria. En este caso se llaman punteaduras .
En las células en crecimiento, las paredes celulares, denominadas primarias, son más delgadas y menos rígidas que en las células diferenciadas.
2.3.4. Orgánulos celulares: mitocondrias, cloroplastos, retículo endoplasmático, complejo de Golgi, lisosomas y vacuolas.
Las células eucariotas presentan un complejo sistema de membranas interno que llega a ocupar la mitad de la célula, el llamado sistema endomembranoso, que divide el contenido celular en compartimentos. En cada uno de ellos se realiza un tipo de reacciones bioquímicas, evitándose así que unas interfieran con otras. Estos compartimentos son: el retículo endoplasmático , el aparato de Golgi, las vacuolas, los lisosomas y los peroxisomas. Además, hay dos tipos de compartimentos, no relacionados estructuralmente con el sistema endomembranoso, que poseen una doble membrana y que son orgánulos productores de energía. Son los cloroplastos y las mitocondrias .
MITOCONDRIAS
Las mitocondrias son los orgánulos de las células eucariotas que se encargan de la obtención de energía mediante la respiración celular, un proceso de oxidación en el que intervienen unas enzimas denominadas ATP-sintetasas. La energía obtenida se guarda en forma de ATP. Aparecen en grandes cantidades en el citoplasma de todas las células eucariotas, tanto de animales como de vegetales, siendo especialmente abundantes en aquellas que por su actividad poseen una elevada demanda de energía bioquímica (ATP). El conjunto de mitocondrias de una célula se denomina condrioma .
Estructura de las mitocondrias
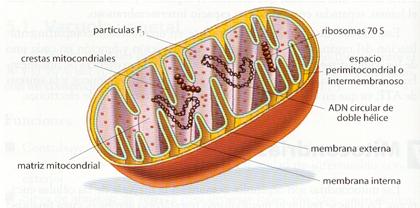
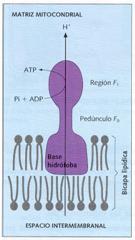
Las mitocondrias son orgánulos polimorfos, pudiendo variar desde formas esféricas hasta alargadas a modo de bastoncillo. Sus dimensiones oscilan entre 1 µ y 4 µ de longitud y 0,3 µ y 0,8 µ de anchura. Presentan una doble membrana: unamembrana mitocondrial externa lisa y una membrana mitocondrial internacon numerosos repliegues internos, denominados crestas mitocondriales. Estas membranas originan dos compartimentos: el espacio intermembranoso, entre las dos membranas, y la matriz, espacio delimitado por la membrana interna.
La membrana mitocondrial externa posee un gran número de proteínas transmembranosas que actúan como canales de penetración. A continuación, se encuentra el espacio intermembranoso, de contenido similar al del citosol. La membrana mitocondrial interna presenta repliegues o crestas que incrementan su superficie y, por tanto, su capacidad metabolizadora. Es bastante impermeable, y presenta un gran número de proteínas de membrana que desarrollan una amplia gama de funciones, destacando las permeasas, los componentes de las cadenas moleculares transportadoras de electrones, entre los que destacan los citocromos, y los complejos enzimáticos formadores de ATP, denominados ATP-sintetasas. Entre sus lípidos de membrana no aparece el colesterol, al igual que en la membrana plasmática bacteriana.
Las ATP-sintetasas están constituidas por tres partes: una base hidrófoba, que se ancla en la membrana, un pedúnculo o región F o, y una esfera de unos 90 Å de diámetro, o región F 1, que es donde se catalizan las reacciones de síntesis de ATP.
En la matriz mitocondrial existe un medio interno rico en enzimas y en el que se lleva a cabo un gran número de reacciones bioquímicas. Esta cámara interna presenta ribosomas mitocondriales o mitorribosomas, similares a los bacterianos, y varias moléculas de ADN mitocondrial, circular y de doble hebra como los bacterianos.
|
|
|
|
Estructura de una mitocondría |
ATP-sintetasa |
Función de las mitocondrias
Su actividad principal es la respiración mitocondrial , cuya última etapa es la cadena respiratoria que se realiza en la membrana interna. En esta se dan los NADH y los FADH2 procedentes de otras vías metabólicas, obteniéndose energía que se almacena en moléculas de ATP. La síntesis de ATP por la ATP-sintetasa se explica mediante quimioósmosis: El NADH y el FADH2 liberan protones (H+) y electrones (e-). Éstos, al pasar por la cadena de proteínas transportadora de electrones que hay en las crestas mitocondriales, ceden energía, que es utilizada para bombear protones H+ fuera de la matriz. Éstos regresan a la matriz a través de las ATP-sintetasas donde la energía del gradiente es utilizada para formar ATP.
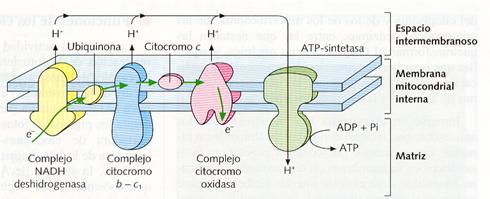
Sector de la membrana interna mitocondrial mostrando la secuencia de proteínas de la cadena respiratoria y la ATP-sintetasa que realizan la fosforilación oxidativa que conduce a la formación de moléculas de ATP
En la matriz mitocondrial se realizan otras vías metabólicas importantes; las principales son:
- El ciclo de Krebs.
- La b-oxidación de los ácidos grasos.
- La biosíntesis de proteínas en los ribosomas.
- La duplicación del ADN mitocondrial.
CLOROPLASTOS
Los cloroplastos son unos orgánulos típicos de las células vegetales que poseen clorofila, por lo que pueden realizar la fotosíntesis, proceso en el que se transforma la energía luminosa en energía química contenida en la molécula de ATP. Por ello, al igual que las mitocondrias, los cloroplastos son orgánulos productores de energía.
Estructura de los cloroplastos
Los cloroplastos son polimorfos y de color verde debido a la presencia del pigmento clorofila. En las algas, las formas son muy diversas; por ejemplo, en el alga Spirogyra sólo hay dos y tienen forma de cinta en espiral. En las plantas superiores, la forma más abundante es la de disco lenticular, aunque también los hay ovoides y esféricos. Miden entre 3 y 19 µ. de diámetro mayor y de 1 y 2 µ de diámetro menor, y suele haber entre 20 y 40 por célula.
Presentan una envoltura constituida por una doble membrana: una membrana plastidial externa y una membrana plastidial interna. La externa es muy permeable, mientras que la interna es casi impermeable, por lo que posee una gran cantidad de permeasas, denominadas proteínas translocadoras. Ambas membranas carecen de clorofila y entre sus lípidos, aligual que en las mitocondrias, no está el colesterol.
En el interior, delimitada por la membrana plastidial interna, hay una cámara que contiene un medio interno, denominado estroma, que posee un elevado número de componentes. Éstos son: ADN plastidial, circular y de doble hélice, como el de las bacterias; plastorribosomas, distintos de los ribosomas del citoplasma y de los mitorribosomas de las mitocondrias; enzimas, entre las que destacan las que transforman el CO2 en materia orgánica, y aquellas que permiten la transcripción, traducción y replicación del ADN; y finalmente, las inclusiones de granos de almidón y las inclusiones lipídicas.
Inmerso en el estroma hay numerosos sáculos aplastados, que reciben el nombre de tilacoides o lamelas, caracterizados por contener pigmentos fotosintéticos en su membrana, la denominada membrana tilacoidal, cuya cavidad interior recibe el nombre de lumen o espaciotilacoidal. Los tilacoides pueden extenderse por todo el estroma, por lo que reciben el nombre de tilacoides de estroma, o pueden ser pequeños, tener forma de disco y presentarse apilados como montones de monedas, los denominados tilacoides de grana, ya que cada montón recibe el nombre de grana. En las membranas de los grana se ubican los sistemas enzimáticos encargados de captar la energía luminosa, efectuar el transporte de electrones y formar ATP.
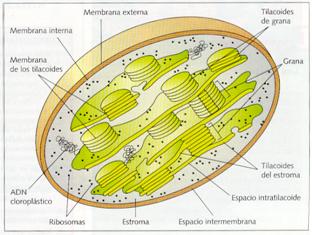
Estructura del cloroplasto.
Funciones de los cloroplastos
La principal actividad de los cloroplastos es la realización de la fotosíntesis, en la que la materia inorgánica es transformada en materia orgánica (fase oscura de la fotosíntesis) utilizando la energía bioquímica (ATP) obtenida a partir de la energía solar, mediante los pigmentos fotosintéticos y la cadena transportadora de electrones de los tilacoides (fase luminosa de la fotosíntesis). Al igual que en las mitocondrias, la síntesis de ATP se realiza mediante la quimioósmosis. Los electrones excitados por la luz, es decir, ricos en energía, al recorrer la cadena de proteínas situadas en la membrana de los tilacoides (cadena transportadora de electrones), van cediendo su energía, que es utilizada para bombear H+ hacia el interior de los tilacoides. Esto origina un gradiente químico de H+, cuya energía es utilizada por las ATP-sintetasas para la formación de ATP.
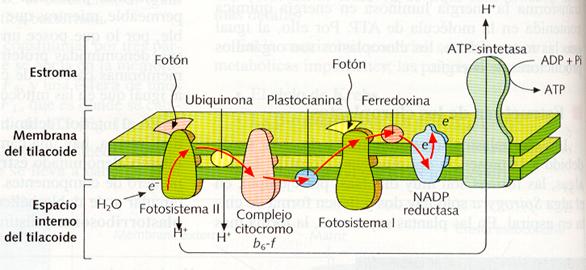
Sector de la membrana de un tilacoide, mostrando las proteínas que realizan el transporte de electrones y a la ATP-sintetasa.
Otras vías metabólicas, que se realizan en el estroma, son la biosíntesis de proteínas y la replicación del ADN.
RETICULO ENDOPLASMATICO
El retículo endoplasmático es un sistema membranoso compuesto por una red de sáculos aplastados o cisternas, sáculos globosos o vesículas, y túbulos sinuosos, que se extienden por todo el citoplasma y que se halla en comunicación con la membrana nuclear externa. Este sistema constituye un único compartimento con un espacio interno que recibe el nombre de lumen. Se distinguen dos clases de retículo endoplasmático: el retículo endoplasmático rugoso o granular (RER), antes denominado ergastoplasma, que posee ribosomas en su cara externa, la llamada cara citoplasmática y el retículo endoplasmático liso o agranular (REL), que carece de ribosomas.
Estructura del retículo endoplasmático rugoso
El retículo endoplasmático rugoso está formado por sáculos aplastados comunicados entre sí. Además, puede presentar vesículas. Se encuentra comunicado con el retículo endoplasmático liso y con la membrana externa de la envoltura nuclear.
De hecho, puede considerarse que la envoltura nuclear es la parte del RER que separa el núcleo del citoplasma. Sus membranas son algo más delgadas que las plasmáticas (50 a 60 Å), y presentan proteínas encargadas de fijar los ribosomas, las riboforinas , y otras que actúan como canales de penetración de las proteínas sintetizadas por estos ribosomas.
Función del retículo endoplasmático rugoso
La función básica del retículo endoplasmático rugoso es la síntesis de proteínas mediante los ribosomas de su membrana, su introducción en el lumen, la glucosilación de las proteínas (que se completará en el aparato de Golgi), y su transporte hacia los orgánulos, donde son utilizadas para constituir membranas.
Este transporte se realiza en el interior de vesículas que se producen en la membrana del retículo endoplasmático rugoso.
|
|
|
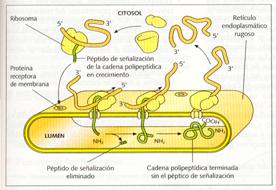
|
Retículo endoplasmático rugoso y su relación con la envoltura nuclear y el retículo endoplasmático liso. |
Síntesis de proteínas en la membrana del retículo endoplasmático rugoso. |
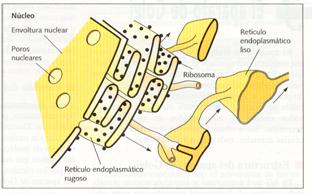
Estructura del retículo endoplasmático liso
El retículo endoplasmático liso está constituido por una red de tubulos unidos al retículo endoplasmático rugoso y que se expande por todo el citoplasma. La membrana del retículo endoplasmático liso posee gran cantidad de enzimas cuya principal actividad es la síntesis de lípidos.
Función del retículo endoplasmático liso
En el REL se sintetizan casi todos los lípidos constituyentes de las membranas: colesterol, fosfolípidos, glucolípidos, etc. Sólo los ácidos grasos se sintetizan en el citosol. Estos lípidos se construyen en el lado citoplasmático de la membrana, desde donde se difunden hacia el interior del retículo liso. Estos lípidos se transportan a otros orgánulos, mediante proteínas de transferencia o por vesículas producidas por medio de las redes de clatrina por gemación. Es decir, intervienen en la síntesis, almacenamiento y transporte de lípidos.
Participa también en procesos de detoxificación , siendo capaz de metabolizar sustancias tóxicas y convertirlas en productos eliminables por las células.
Por último, interviene en la conducción de impulsos nerviosos para la contracción del músculo estriado.
COMPLEJO DE GOLGI
El aparato de Golgi (AG) forma parte del sistema membranoso celular. Fue descubierto por C. Golgi en 1898. Está formado por uno o varios dictiosomas (agrupación en paralelo de cuatro a ocho sáculos discoidales denominados cisternas), acompañados de vesículas de secreción. Suele situarse próximo al núcleo, y, en las células animales, rodeando a los centríolos.
Estructura del aparato de Golgi.
El AG está estructural y fisiológicamente polarizado, ya que el dictiosoma presenta una cara cis , próxima al RER, generalmente convexa, constituida por sáculos de menor diámetro y de membrana más fina, y una cara trans, próxima a la membrana citoplasmática, generalmente cóncava, y caracterizada por presentar cisternas de gran tamaño, de membrana más gruesa y de aspecto reticular. La cara cis o de formación recibe vesículas (vesículas de transición) procedentes de la envoltura nuclear y del retículo endoplasmático, que alimentan al aparato de Golgi. El contenido molecular del dictiosoma va avanzando hacia la cara trans o de maduración. Esta progresión se realiza de cisterna a cisterna, mediante pequeñas vesículas (vesículas intercisternas) y, una vez que llega a la cara trans, es concentrado y acumulado en el interior de unas vesículas mucho mayores que las anteriores (vesículas de secreción). Éstas pueden actuar como lisosomas si contienen enzimas digestivas o pueden dirigirse hacia la membrana plasmática donde pueden verter su contenido al medio externo (exocitosis ) y además soldarse a ella y, así, hacerla crecer o regenerarse.
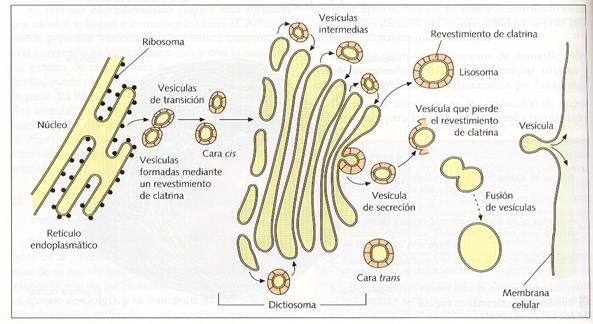
Origen del aparato de Golgi y los lisosomas.
Función del aparato de Golgi
Las membranas del AG son ricas en proteínas tipo de enzimático, por lo que pueden realizar múltiples funciones. Desempeña el papel de organizador de la circulación molecular de la célula. Por él pasan gran número de moléculas procedentes del retículo endoplasmático que sufren una maduración en su recorrido por los sáculos del dictiosoma.
Entre las funciones del aparato de Golgi destacan:
- Transporte, maduración, acumulación y secreción de proteínas procedentes del retículo endoplasmático. Así, muchas proteínas varían su estructura o alteran las secuencias de aminoácidos haciéndose activas. Posteriormente son concentradas y pasan al interior de vesículas de secreción.
- Glucosilación de lípidos y proteínas, mediante la unión a éstos de cadenas de oligosacáridos, dando lugar a glucolípidos o glucoproteínas de membrana, o de secreción.
- Síntesis de proteoglucanos (mucopolisacáridos), que son parte esencial de la matriz extracelular, y de los glúcidos constitutivos de la pared celular vegetal (pectina, hemicelulosa y celulosa).
LISOSOMAS
Los lisosomas son vesículas procedentes del aparato de Golgi que contienen enzimas digestivas. Éstas son hidrolasas ácidas (fosfatasa ácida, glucosidasas, lipasas, proteasa, ADNasa, etc.) que se forman en el retículo endoplasmático rugoso, pasan al aparato de Golgi, en donde se activan y se concentran, y que se acumulan en el interior de los lisosomas.
Estructura de los lisosomas
Los lisosomas poseen una membrana plasmática conlas proteínas de su cara interna muy glucosiladas. Estas glucoproteínas impiden que las enzimas hidrolasas ataquen a la propia membrana del lisosoma.
Función de los lisosomas
Los lisosomas realizan la digestión de materia orgánica. La principal enzima digestiva es la fosfatasaácida , capaz de romper los enlaces fosfoestéricos y liberar grupos fosfato. Para el buen funcionamiento de estas enzimas, los lisosomas necesitan mantener un pH entre 3 y 6; por tanto, introducen protones (H+) en su interior mediante gasto de ATP.
La digestión puede ser extracelular , cuando los lisosomas vierten sus enzimas al exterior, o intracelular, cuando se unen a una vacuola que contiene la materia a digerir. Se utiliza el término de lisosoma primariopara referirse a los que sólo poseen en su interior enzimas digestivas y el término lisosoma secundariopara aquellos que, por haberse unido a una vacuola con materia orgánica, contienen sustratos en vía de digestión. Los lisosomas secundarios reciben el nombre de vacuolas digestivas o heterofágicas, cuando el sustrato procede del exterior por fagocitosis o pinocitosis, o de vacuolas autofágicas, cuando procede del interior, por ejemplo, con moléculas u orgánulos propios, que previamente han sido envueltos por cisternas del retículo endoplasmático.
Esquema de la acción de los lisosomas en heterofagia y en autofagia.
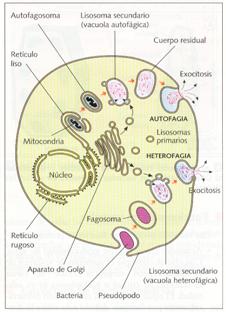
Lisosomas especiales son el acrosoma de los espermatozoides y los granos de aleurona de las semillas. El acrosoma es unlisosoma primario en el que se almacenan enzimas capaces de digerir las membranas foliculares del óvulo, para permitir el paso del espermatozoide y la fecundación. Los granos de aleurona son lisosomas secundarios en donde se almacenan proteínas que, debido a la pérdida de agua, se encuentran en estado cristalino, hasta que al plantarse y absorberse agua se activan las enzimas y se inicia la digestión de las mismas, con lo que empieza la germinación de la semilla.
Los peroxisomas y los glioxisomas.
Los peroxisomas son orgánulos parecidos a los lisosomas, pero en vez de contener enzimas hidrolasas contienen enzimas oxidasas, entre las que destacan la peroxidasa y la catalasa.
Los glioxisomas son una clase de peroxisomas que sólo se encuentran en las células de los vegetales.
Una función de los peroxisomas es la detoxificación, proceso que elimina sustancias tóxicas oxidándolas. Así se degradan, en las células hepáticas, el etanol de las bebidas alcohólicas y otras sustancias tóxicas como el metanol, el ácido fórmico, etc.
VACUOLAS
Las vacuolas son vesículas constituidas por una membrana plasmática, y cuyo interior es predominantemente acuoso. Cuando en el contenido hay otro tipo de sustancias predominantes se habla de inclusiones.
Estructura de las vacuolas
Las vacuolas se forman a partir del retículo endoplasmático, del aparato de Golgi o de invaginaciones de la membrana citoplasmática. Las vacuolas de las células animales, que ya se han descrito al estudiar el RE y el AG, suelen ser pequeñas, y actualmente se denominan vesículas. Las vacuolas de las células vegetales suelen ser muy grandes, y sí reciben el nombre de vacuolas. Suele haber una o dos en cada célula. La membrana recibe el nombre de tonoplasto. Se forman mediante la unión de vesículas derivadas del RE y del AG, que suelen ocupar un 5% del volumen celular. A medida que la célula vegetal joven madura, las vacuolas crecen, llegando a ocupar un 50 %, y en ocasiones hasta un 90 %, de la célula vegetal madura. El conjunto de vacuolas de una célula vegetal recibe el nombre de vacuoma.
Funciones de las vacuolas
Las vacuolas poseen tres funciones principales:
- Acumular en su interior gran cantidad de agua. Con ello se consigue el aumento de volumen de la célula vegetal -turgencia celular- sin variar la cantidad de citosol o hialoplasma ni su salinidad. El agua, que realiza una función estructural, entra por ósmosis debido a la elevada concentración de sustancias que hay en las vesículas iniciales.
- Sirven de almacén de muchas sustancias. Unas son reservas energéticas elaboradas por la por la propia célula, como proteínas, otras son productos de desecho que resultarían perjudiciales si estuvieran en el citosol, otras son sustancias con funciones específicas, por ejemplo, los antocianósidos responsables de los colores de los pétalos, o los alcaloides venenosos para repeler los animales herbívoros, y otras son sustancias con función esquelética, como los cristales de carbonato cálcico y oxalato cálcico.
- Son medio de transporte entre orgánulos del sistema endomembranoso y entre éstos y el medio externo. Lo realizan las llamadas vesículas del RE y del AG.
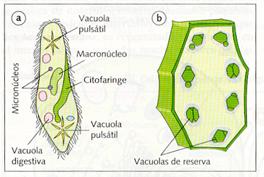
Por último, en las células animales se conocen dos tipos especiales de vacuolas: unas con función nutritiva, como las vacuolas fagocíticas y las pinocíticas, y otras con función reguladora de la presión osmótica; éstas son las vacuolas pulsátiles de los protozoos ciliados, que expulsan agua al exterior de una forma rápida, si la diferencia de presión es grande, o de una forma lenta, si los medios son isotónicos.
Entre las inclusiones, las más frecuentes son las inclusiones lipídicas , de aspecto muy refringente, que pueden contener lípidos de reserva o gotas de aceite, que por oxidación dan origen a las resinas y a los depósitos de látex, sustancia de la cual deriva el caucho natural.
|
a) Paramecio con vacuolas pulsátiles y digestivas. b) Célula vegetal con vacuolas de reserva de sustancias oleosas.
|
|
2.3.5. Núcleo: envoltura nuclear, nucleoplasma, cromatina y nucleolo.
Se trata de un corpúsculo relativamente grande que fue observado por Brown en 1831, aunque se tenía constancia de su existencia desde unas observaciones de Leeuwenhoek en 1691. Está presente en prácticamente todas las células eucariotas, aunque existen excepciones, como los glóbulos rojos. Normalmente hay un núcleo por célula; si bien existen células con dos núcleos o binucleadas, como los paramecios y algunos hongos, y masas protoplásmicas plurinucleadas, como son los plasmodios y los sincitios.
Su forma es normalmente esférica, con un diámetro que oscila entre los 5 y los 25 nm, aunque también puede presentarse con forma ovoide, en células epiteliales prismáticas, forma alargada, en células fusiformes, irregular, en células hiperactivas, e incluso polilobulado, en los granulocitos. Su posición es variable, central, en la mayoría de las células animales, y lateralizado en células vegetales y algunas animales como las adiposas y las mucosas.
Alberga la información genética y, por tanto, controla la actividad celular. Se piensa que una de las razones de la aparición del núcleo en la evolución desde procariontes a eucariontes fue, precisamente, proteger al ADN frente a las tensiones de los filamentos del citoesqueleto.
Puede aparecer en dos estados bien diferentes: en reposo o núcleo interfásico, que será el objeto de nuestro estudio, y en división o núcleo mitótico, este último cuando la célula entra en mitosis. En el núcleo en reposo pueden distinguirse claramente cuatro estructuras diferentes: membrana nuclear, jugo nuclear, cromatina y nucleolos.
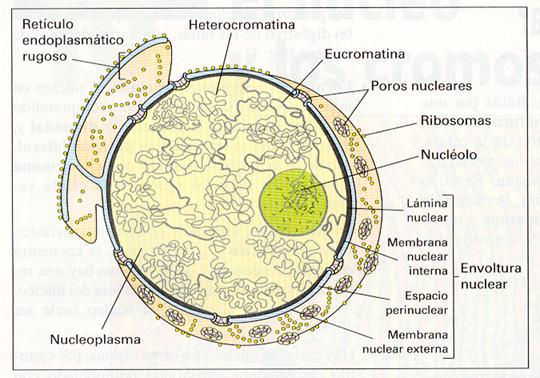
Ultraestructura del núcleo.
ENVOLTURA NUCLEAR.
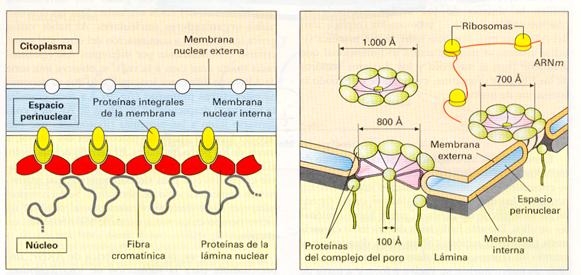
Consiste en una doble membrana sin pliegues que presenta cierta continuidad con la del retículo endoplásmico. Se caracteriza por poseer numerosos poros como resultado de la fusión de ambas membranas en determinados puntos denominados complejos de poro. La membrana interna tiene asociada un material denso a los electrones denominado lámina fibrosa o corteza nuclear; la membrana externa, por su parte, puede tener adheridos ribosomas, de manera similar a lo que ocurre en el RER.
Los poros, cuyo número y distribución dependen del tipo celular, siendo el primero mayor cuanto más activa es la célula. Resulta característica su apariencia poligonal en células germinales, apreciándose una serie de subunidades dispuestas en círculo. La función de los poros es permitir el transporte bidireccional de sustancias.
|
|
|
Envoltura nuclear. Complejo del poro nuclear. |
NUCLEOPLASMA
También denominado jugo nuclear o carioplasma, se trata del contenido semilíquido del núcleo y su composición es semejante a la del hialoplasma, lo que es lógico si se piensa que los poros permiten plena libertad de movimiento de las sustancias entre núcleo y citoplasma.
Incluye gran cantidad de proteínas y enzimas involucradas en la replicación del ADN, así como en la transcripción del ARN y su empaquetamiento para el traslado al citoplasma.
CROMATINA.
En las células eucarióticas cada molécula de ADN está densamente empaquetada alrededor de proteínas estructurales formando la cromatina. La razón del empaquetamiento es doble, ya que además de conseguir una drástica reducción en el gran tamaño del material genético, se piensa también que las diferencias observadas en el patrón del mismo guardan relación con la actividad de los genes.
Las proteínas presentes en la cromatina se clasifican en histonas y no histonas. Las histonas son exclusivas de eucariontes y se encuentran en una cantidad igual a la de ADN. Son proteínas de pequeño tamaño que contienen una alta proporción de aminoácidos cargados positivamente (lisina y arginina constituyen hasta el 24% del polipéptido), lo que les permite unirse fuertemente con el ADN, puesto que éste tiene carga negativa. Se piensa que sólo en contadas ocasiones el ADN se disocia de sus histonas, por lo que se cree que deben jugar cierto papel en todas las reacciones en las que el material genético se ve involucrado.
Se distinguen dos grupos de histonas:
- Histonas nucleosomales (H2A, H2B, H3 y H4), responsables del empaquetamiento del ADN en los denominados nucleosomas y que son similares en la mayoría de las células, lo que muestra su alta conservación a lo largo de la filogenia.
- Histonas H1, que presenta formas específicas para diferentes tipos celulares, lo que indica que su conservación evolutiva ha sido mínima y que, como se verá más abajo, están implicadas en el apilamiento o condensación de los nucleosomas.
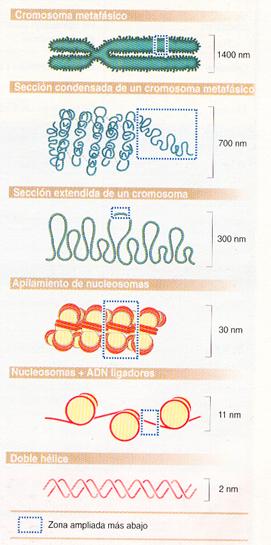
El concepto de nucleosoma fue propuesto en 1974 por R.D. Kornberg y se le considera la unidad de empaquetamiento del ADN. Cuando el material genético es descondensado presenta una típica apariencia de collar de perlas en micrografías electrónicas, apreciándose los nucleosomas enlazados por segmentos de ADN denominados ADN ligador . El nucleosoma consiste en un octámero de histonas alrededor del cual se enrolla la doble hélice del ADN y aparece como una estructura cilíndrica de 11 nm de diámetro. Cada nucleosoma permite el enrollamiento de aproximadamente 200 nucleótidos y se encuentra unido con el siguiente nucleosoma por un ADN ligador de hasta 80 nucleótidos.
|
|
|
Estructura de la cromatina en “collar de perlas”. |
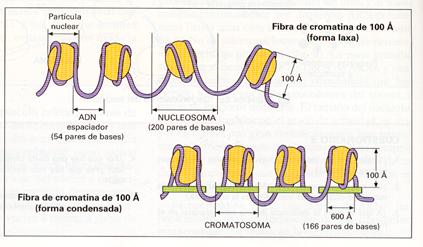
Pero los nucleosomas por sí solos no pueden explicar el alto grado de empaquetamiento del material genético. Así pues, los nucleosomas se van a apilar unos con otros gracias a las histonas Hl, de forma que la cromatina se observa, en lo que es su estructura natural, como una fibra de unos 30 nm de diámetro. En ocasiones, la homogeneidad puede romperse y las fibras presentan bloques de nucleosomas engarzados por proteínas.
Pero el grado de condensación de la cromatina puede ser aún mayor, por lo que puede hablarse, por tanto, de dos tipos de cromatina en el núcleo interfásico: la eucromatina, que sería el material genético en su estado más difuso, y la heterocromatina, cuando presenta un mayor grado de condensación, como es el caso de los cromosomas metafásicos o regiones concretas de un cromosoma que permanecen condensadas también en la interfase. La heterocromatina puede ser, a su vez, constitutiva, pues aparece condensada en todas las células del organismo y es genéticamente inactiva, y facultativa, que representa bien a cromosomas, bien a regiones cromosómicas que normalmente son inactivadas, permanentemente o no, según el tipo celular o el período de desarrollo.
El mayor grado de condensación del material genético se consigue cuando la célula va a dividirse y se habla entonces de cromosomas.
|
|
|
Diferentes grados de compactación de la cromatina , desde el mínimo representado por la doble hélice hasta el máximo, que corresponde al cromosoma metafásico. |
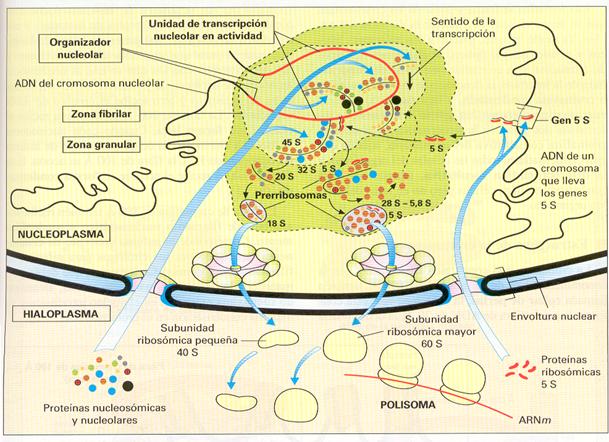
NUCLEOLO
Región esferoidal del núcleo que presenta una alta concentración de ARN y proteínas y que aparece en un lugar específico del mismo, en número de uno o dos. Como se ha mencionado anteriormente, se relacionan con constricciones secundarias de algunos cromosomas conocidas como regiones organizadoras del nucleolo (NOR) u organizadores nucleolares. Su tamaño depende del grado de actividad de la célula, ya que su función es la síntesis de los ARN y su procesado y empaquetamiento para formar las subunidades de los ribosomas.
Al microscopio electrónico el nucleolo presenta una doble composición. Un componente nuclear que consiste en la cromatina asociada al nucleolo y un componente propio del nucleolo que puede ser, a su vez, de dos tipos: fibrilar y granular. El fibrilar se corresponde con moléculas de ARN asociadas a proteínas, mientras que el granular consiste en subunidades ribosomales en proceso de maduración.
|
|
|
Esquema del funcionamiento del nucleolo. |