1.7. Ácidos nucleicos.
1.7. Ácidos nucleicos.
El descubrimiento de la sustancia que luego se llamó ADN fue realizado en 1869 por Miescher, quien comenzó el estudio con leucocitos obtenidos del pus de los vendajes y después con espermatozoides de salmón. Obtuvo una sustancia que contenía C, H, O, N y un alto porcentaje de P, a la que llamó nucleína, por proceder del núcleo de dichas células. Años más tarde fragmentó esta nucleína y separó el componente proteico, de naturaleza básica. El grupo prostético reveló su carácter ácido y se le llamó ácido nucleico.
La determinación de la estructura de la nucleína fue muy lenta y, en los años 30, Kossel demostró que las nucleínas no eran simples mezclas de proteínas y ácido nucleico, sino que eran auténticos complejos supramoleculares, conocidos por los histólogos como cromatina. La cromatina, durante la etapa de división celular se organiza en cromosomas. A finales del siglo XIX se había reconocido que los cromosomas eran los portadores de la información hereditaria. Pero la información podía residir en las proteínas o en el ADN. La evidencia de que es el ADN de estos cromosomas la molécula que contiene la citada información no se obtuvo hasta 1944.
Los estudios químicos de Kossel establecieron que la subunidad repetitiva del ADN es un nucleótido que contiene azúcar, un grupo fosfato y una de las cuatro bases nitrogenadas heterocíclicas.
Trabajos posteriores descubrieron cómo están unidos estos componentes. En los años 40, Chargaff demostró que el número de bases de adenina es igual al número de bases de timina, y el número de bases de citosina coincide con el de guanina. Pero no fue hasta 1953 cuando James Watson y Francis Crick descubrieron la estructura tridimensional del ADN, basándose en la equivalencia de bases de Chargaff y en los estudios de difracción de rayos X sobre fibras de ADN obtenidos por Rosalind Franklin y Maurice Wilkins.
1.7.1. Concepto e importancia biológica.
Los ácidos nucleicos son biomoléculas orgánicas compuestas siempre de carbono, hidrógeno, oxígeno, nitrógeno y fósforo. Se definen químicamente como polinucleótidos, ya que están formados por la repetición de unidades moleculares llamadas nucleótidos.
Existen dos tipos de ácidos nucleicos: el ácido desoxirribonucleico (ADN) y el ácido ribonucleico (ARN).
Estas moléculas que poseen todos los organismos, dirigen y controlan la síntesis de proteínas, proporcionando la información que determina su especificidad y características biológicas y contienen las instrucciones necesarias para realizar los procesos vitales y son los responsables de todas las funciones básicas de los seres vivos. Podría decirse que lo que un organismo es o puede llegar a ser, en términos biológicos, aparece “programado” en estas moléculas. O dicho de otro modo, su función está relacionada con el almacenamiento y la transmisión de la información genética constituyendo la base molecular de la herencia.
1.7.2. Nucleótidos. Enlace fosfodíéster. Funciones de los nucleótidos.
NUCLEOTIDOS
Los nucleótidos son las unidades que forman los ácidos nucleicos. En este sentido, equivalen a los monómeros que constituyen las otras macromoléculas biológicas.
Cada nucleótido es una molécula relativamente compleja, compuesta por la unión de tres unidades: un monosacárido (una pentosa), una base nitrogenada y uno o varios grupos fosfato. Tanto la base nitrogenada como los grupos fosfato se encuentran unidos a la pentosa.
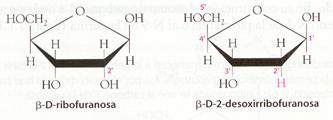
La pentosa siempre es una aldopentosa, ya sea b-D-ribofuranosa, y en este caso el nucleótido se denomina ribonucleótido, o b-D-2-desoxirribofuranosa, constituyente de los desoxirribonucleótidos (el prefijo desoxi- indica que la pentosa carece de un átomo de oxígeno).
La base nitrogenada puede ser de dos tipos: púrica, derivada de la purina o pirimidínica, derivada del anillo de la pirimidina.
La presencia de los átomos de nitrógeno comunica, en ambos casos, carácter básico a estos compuestos.
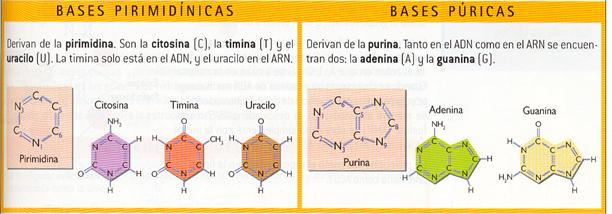
La existencia de distintos radicales hace que puedan aparecer varias bases nitrogenadas; concretamente, las presentes en los ácidos nucleicos son dos bases derivadas de la purina, la adenina (A) y la guanina (O), y tres derivadas de la pirimidina, la citosina (C), la timina (T) y el uracilo (U).
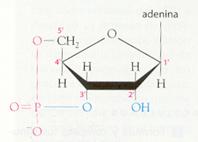
La porción constituida por la pentosa y la base nitrogenada se denomina nucleósido . En su constitución, el átomo de carbono C-1 de la pentosa se une al nitrógeno N-1 de la pirimidina o al N-9 de la purina.
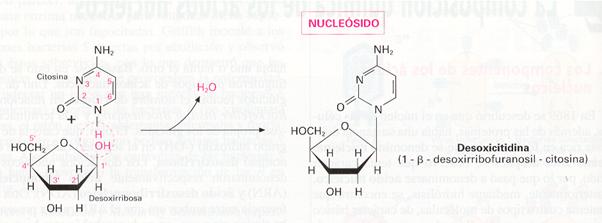
Para distinguir qué átomos pertenecen a la pentosa y cuáles a la base se emplean en la primera números con apóstrofe. Así, por ejemplo, diríamos que para constituir un nucleósido, la base nitrogenada se une al carbono l' de la pentosa.
Como existen dos clases de pentosas y cinco bases nitrogenadas, pueden aparecer diez tipos de nucleósidos diferentes. Para nombrarlos, se antepone al nombre del nucleósido un prefijo que indica la base nitrogenada (cit-, tim-, ur-, aden-, guan-,) y se añade la terminación -idina, si la base es pirimidínica, y -osina, si es púrica.
Si la pentosa es la desoxirribosa, se coloca el prefijo desoxi, delante del nombre. Si es la ribosa, no se antepone ningún prefijo. Por ejemplo, los cuatro nucleósidos del ADN se llaman desoxiadenosina, desoxiguanosina, desoxitimidina y desoxicitidina.
La unión de uno o varios grupos fosfato al carbono 3' o al carbono 5' de la pentosa da lugar al nucleótido completo. Su nomenclatura es sencilla: se elimina la última letra a del nombre del nucleósido y se indica a continuación el lugar de unión a la pentosa y el número de fosfatos unidos. Si no se indica numeración, se sobreentiende que la unión se realiza con el carbono 5'.
Cuando existe más de un grupo fosfato, se unen en cadena, uno a continuación de otro.
En algunas representaciones se observa que los grupos -OH unidos a los átomos de P figuran como -O-. Esto es debido a que a los pH biológicos habituales esos grupos se encuentran ionizados.
Con frecuencia se emplean abreviaturas para nombrar a los nucleótidos. Así GTP indica guanosín trifosfato.
ENLACE FOSFODIESTER.
La existencia de grupos hidroxilo, tanto en la pentosa como en el fosfato, permite la unión de los nucleótidos mediante la formación de enlaces entre ambos grupos.
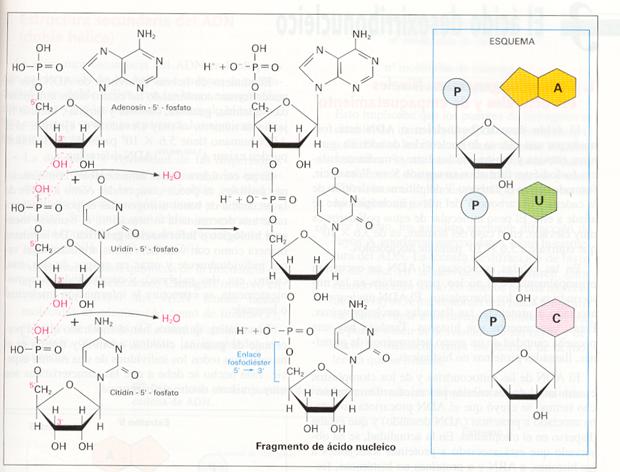
Formación del enlace fosfodíéster, constituyendo un trinucleótido.
La unión es una esterificación que se realiza entre el grupo fosfato situado en posición 5' de un nucleótido y el grupo hidroxilo que se encuentra en el carbono 3' de otro nucleótido.
Se trata, por tanto, de una condensación, en la que se obtiene un compuesto denominado dinucleótido y se libera una molécula de agua. El nuevo enlace, de tipo éster fosfórico, se denomina enlace fosfodíéster, o enlace nucleotídico. La hidrólisis del dinucleótido libera los dos mononucleótidos.
Dado que existen aún grupos hidroxilo libres, el dinucleótido se puede unir a más nucleótidos y formar trinucleótidos, tetranucleótidos, etc. La unión de cientos o miles de cadenas de nucleótidos constituye los ácidos nucleicos o polinucleótidos, moléculas gigantescas con una masa molecular muy elevada.
Como la unión entre los nucleótidos se produce entre pentosas y fosfatos, las bases nitrogenadas no participan en los enlaces. Se establecen, por tanto, largas cadenas sin ramificar formadas por la secuencia pentosa- fosfato-pentosa- fosfato…, en la que las bases nitrogenadas quedan «colgando» lateralmente de las pentosas. La secuencia de estas bases nitrogenadas es la que proporciona la especificidad a una cadena polinucleotídica determinada.
En todos los polinucleótidos existe un extremo (denominado extremo 3') con una pentosa con el grupo -OH del carbono 3' libre, y otro extremo (llamado extremo 5'), donde se localiza un grupo fosfato libre unido al carbono 5' de otra pentosa. Es decir, ambos extremos son distintos.
Se suele emplear una notación simplificada para indicar la composición de un polinucleótido, en la que únicamente se señala la secuencia de bases nitrogenadas y los extremos 5' y 3'.
FUNCIONES DE LOS NUCLEOTIDOS.
Los nucleótidos son moléculas que poseen un gran interés biológico, ya que, además de constituir los ácidos nucleicos (función estructural), llevan a cabo algunas funciones básicas para los seres vivos, cuando se encuentran libres en la célula. Entre las principales funciones tenemos:
Moléculas acumuladoras y donantes de energía.
Ciertas reacciones bioquímicas propias de los seres vivos tienen como finalidad la producción de energía. Si esta energía se desprendiera libremente apenas tendría utilidad para el organismo. Resulta más eficaz disponer de un sistema capaz de acumular la energía liberada, de manera que pueda ser utilizada con posterioridad en la cantidad y en el momento precisos. Algunos nucleótidos (fundamentalmente de adenosina y, en menor medida, también de guanosina) con más de un grupo fosfato desempeñan esta función.
Cuando existe energía disponible, una molécula de adenosín difosfato (ADP) la emplea en unir un tercer grupo fosfato a los otros dos para obtener adenosín trifosfato (ATP).
El enlace así constituido es altamente energético, lo que quiere decir que para su formación se requiere una cantidad considerable de energía (7 kcal/mol). Resulta evidente que la rotura de este enlace liberará la misma cantidad de energía.
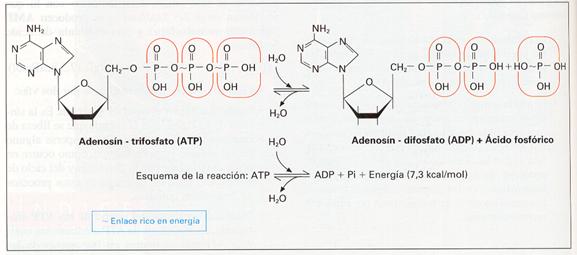
De esta manera, el sistema ADP/ATP constituye una forma eficaz de «guardar» la energía liberada en reacciones biológicas exotérmicas.
Si existe energía disponible en abundancia se formarán muchas moléculas de ATP a partir de ADP. La proporción ATP/ADP será en este caso alta. Por el contrario, el consumo de energía en gran cantidad por parte de la célula disminuirá los niveles de ATP y aumentará los de ADP. La proporción ATP/ADP será más baja.
Es habitual representar los enlaces de alta energía que existen entre los grupos fosfato con el símbolo (~), en lugar de la notación usual (-) de los enlaces.
Moléculas con función coenzimática:
Ciertos dinucleótidos intervienen como coenzimas en algunas reacciones enzimáticas importantes. Este es el caso de:
· Nicotinamín adenín dinucleótido (NAD+). Es un derivado de la vitamina PP o nicotinamida.
· Nicotinamín adenín dinucleótido fosfato (NADP+). Su fórmula es igual a la del NAD+, pero lleva un grupo fosfato en el carbono 2' del nucleótido de adenina.
· Flavín adenín dinucleótido (FAD). Es un derivado de la riboflavina o vitamina B2.
Las tres coenzimas tienen una acción semejante. Participan en reacciones de deshidrogenación, fundamentales en el catabolismo celular. En estas deshidrogenaciones toman H+ y electrones de algunas moléculas y quedan como NADH, NADPH y FADH2, respectivamente. Estas coenzimas, en estado reducido, pueden ceder fácilmente electrones y H+ a otras moléculas.
- El coenzima A que actúa como transportador de grupos acilo (CH3-CO) incluye un nucleótido (ADP) en su molécula.
Mensajeros intracelulares:
La molécula de adenosín monofosfato cíclico (AMPc) desempeña un papel clave en el desencadenamiento de las respuestas de la célula ante las informaciones que recibe del medio extracelular. La unión de moléculas mensajeras (hormonas, neurotransmisores), procedentes de otros lugares, a ciertos receptores específicos de la membrana plasmática provocan la activación de la enzima adenil ciclasa, la cual lleva a cabo la siguiente reacción:
|
|
ATP adenil ciclasa AMPC + P−P |
|
El AMPC sintetizado permite la ejecución de varios procesos bioquímicos que, en última instancia, originan la respuesta celular. Actúa así como mediador entre la información externa y la respuesta final. Por esta razón, también se denomina segundo mensajero .
Molécula de adenosín monofosfato cíclico (AMPc).
1.7.3. Tipos de ácidos nucleicos. Estructura, localización y funciones.
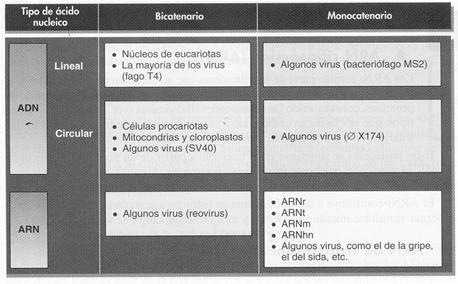
Los ácidos nucleicos son polinucleótidos, o sea, polímeros de nucleótidos. Se distinguen dos tipos: los constituidos por cadenas de desoxirribonucleótidos o ácidos desoxirrionucleicos, ADN (en inglés DNA); y los formados por unidades de ribonucleótidos, ácidos ribonucleicos, ARN (en inglés RNA). La primera diferencia queda clara en los nombres: el ADN contendrá la pentosa desoxirribosa y la ribosa formará parte del ARN.
En un esquema general se observará que el esqueleto del polinucleótido está formado por moléculas de pentosa que se alternan con restos de fosfato, de forma que la cadena está constituida por unidades de pentosa-fosfato enlazadas de la siguiente forma:
...3'-pentosa-5'-fosfato-3'-pentosa-5'-. ..
Por tanto, la cadena tiene una orientación. No es igual un extremo al otro y, en consecuencia, habrá que tenerlo en cuenta a la hora de representar cualquier molécula de ácido nucleico.
Cada resto de pentosa está unido por el carbono l' a una base nitrogenada. Aquí está la segunda diferencia entre los dos tipos de ácidos nucleicos:
- El ADN contiene las bases adenina, guanina, citosina y timina.
- El ARN contiene adenina, guanina, citosina y uracilo.
Así, la timina está ausente en el ARN y el uracilo en el ADN.
Como la cadena es lineal, las bases nitrogenadas seguirán un orden o secuencia. Los ácidos nucleicos son moléculas informativas. En fa información está la clave de su función.
ADN.
De igual forma que las proteínas, el ADN posee diferentes niveles de complejidad estructural. Presenta fundamentalmente estructura primaria y secundaria, aunque asociada o no proteínas nucleares adopta estructuras superenrolladas o empaquetadas que equivaldrían a una estructura terciaria.
Estructura primaria
Es la secuencia de nucleótidos, de estructura y dimensiones conocidas, unidos por enlaces fosfodiéster. Los enlaces fosfodiéster se establecen entre el radical fosfato situado en el carbono 5' de un nucleótido y el radical hidroxilo (-OH) del carbono 3' del siguiente nucleótido (enlaces 5'-3').
Una cadena de ADN presenta dos extremos libres: e unido al grupo fosfato, y el 3', unido a un hidroxilo.
Cada cadena se diferencia de otra por su tamaño, su composición y por su secuencia de bases, que indica el orden en que A, T, C y G se sitúan en la cadena de ADN Cuando se representa una cadena de ADN normalmente solo se indica la secuencia de desoxirribonucleótidos de manera abreviada. Así, la desoxiadenosín-5' monofosfato (AMP) se abreviaría simplemente con la inicial de la base que contiene: A
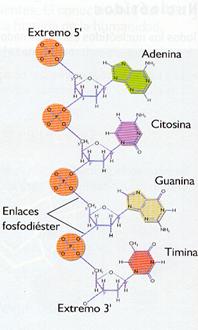
Este polinucleótido se abreviaría como ACGT.
Estructura secundaria
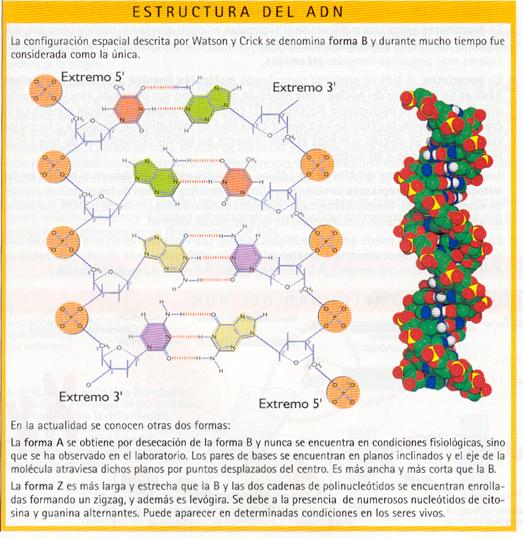
La secuencia polinucleotídica se dispone en el espacio en forma de una doble hélice, según la estructura propuesta por James Watson y Francis Crick en 1953. Dos descubrimientos previos abrieron el camino a este modelo de estructura secundaria del ADN aceptado en la actualidad.
En 1950, Erwin Chargaff, tras estudiar gran cantidad de muestras de ADN pertenecientes a diversas especies de organismos, observó que siempre existía la misma cantidad de bases nitrogenadas púricas y pirimidínicas. Descubrió, además, que el número de adeninas siempre es igual al de timinas, y el de guaninas, al de citosinas. Estos resultados constituyen la denominada ley de equivalencia de bases de Chargaff.
Por otra parte, en esta misma época, Rosalind Franklin y Maurice Wilkins aplicaron el método de difracción de rayos X al ADN y dedujeron que esta molécula posee una estructura helicoidal con dos periodicidades, luna cada 0,34 nm y otra cada 3,4 nm.
A partir de estos datos, Watson y Crick elaboraron su modelo de estructura tridimensional del ADN, que presenta las siguientes características:
- El ADN está constituido por dos cadenas polinucleotídicas unidas entre sí en toda su longitud.
- Las dos cadenas son antiparalelas, lo que significa que el extremo 3' de una de ellas se enfrenta con el extremo 5' de la otra.
- La unión entre las cadenas se realiza por medio de puentes de hidrógeno entre las bases nitrogenadas de ambas: concretamente, la adenina forma dos de estos puentes con la timina y la guanina tres con la citosina. Resulta evidente que las dos cadenas no son idénticas, sino complementarias, ya que una de ellas tiene la secuencia de bases complementaria de la otra. (Si se conoce una secuencia es posible deducir inmediatamente la complementaria.).
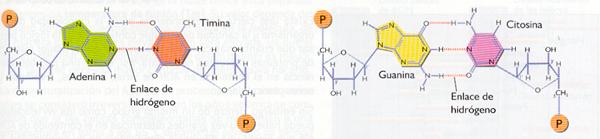
Complementariedad de las bases.
- Las dos cadenas están enrolladas en espiral formando una doble hélice alrededor de un eje imaginario.
- Las bases nitrogenadas quedan en el interior de la doble hélice, mientras que los esqueletos pentosa-fosfato se sitúan en la parte externa. De esta forma, las cargas negativas de los grupos fosfato se unen a las cargas positivas de cationes o de otras moléculas presentes en el medio, estabilizando la estructura.
- Los planos de las bases nitrogenadas enfrentadas son paralelos entre sí y perpendiculares al eje de la hélice.
- El enrollamiento de la doble hélice es plectonémico, es decir, las cadenas no se pueden separar sin desenrollarlas.
- La doble hélice es dextrógira: el enrollamiento gira en el sentido de las agujas del reloj.
- La anchura de la hélice es de 2 nm, la longitud de cada vuelta es de 3,4 nm y cada 0,34 nm se encuentra un par de bases complementarias. Puede deducirse, por tanto, que existen 10 pares de nucleótidos por cada vuelta.
|
|
|
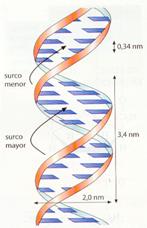
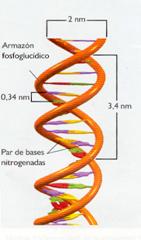
Esquemas de la doble hélice de ADN.
La molécula de ADN es muy estable en condiciones fisiológicas normales, debido a los numerosos enlaces de hidrógeno entre las bases nitrogenadas ya las interacciones hidrofóbicas entre los anillos aromáticos y los radicales -CH3 de las bases nitrogenadas. Sin embargo, la estructura en doble hélice del ADN se puede perder, separándose las dos hebras, cuando se alteran las condiciones de pH (se incrementa por encima de 13) o se calienta a temperaturas en torno a los 100 °C. Luego uno de los agentes físicos que puede producir la separación de las dos hebras de la doble hélice de ADN es la temperatura.
Al fenómeno de separación de las dos hebras de la doble hélice de ADN se denomina desnaturalización.
Los enlaces que se ven afectados y cuya rotura por agentes físicos provoca la separación de las dos hebras de la doble hélice de ADN son, fundamentalmente, los enlaces de hidrógeno que existen entre las bases nitrogenadas: dos enlaces de hidrógeno entre la adenina y la timina, y tres entre la guanina y la citosina.
La desnaturalización del ADN es un proceso que durante varios años se consideró irreversible, pero en los años sesenta se comprobó que si las hebras complementarias se mantenían a una temperatura en torno a los 65 °C durante un periodo de tiempo prolongado, formaban una nueva hélice con total funcionalidad. La desnaturalización es, por tanto, reversible y el proceso inverso, por el que se recupera la doble hélice, se denomina renaturalización o hibridación del ADN.

Desnaturalización y renaturalización de ADN
Estructura terciaria o empaquetamiento.
Las moléculas de ADN tienen un carácter ácido, porque cada nucleótido proporciona una carga electronegativa que procede del grupo fosfato. Por tanto, una molécula con miles o millones de nucleótidos tendrá dos cargas negativas por cada par de bases. Todas estas cargas tienden a repelerse entre sí. En disolución, muchas de ellas se neutralizan con cationes monovalentes, como iones Na+, pero estas moléculas tienden a "esponjarse" y ocupar mucho espacio.
Para codificar toda la información que se requiere en el desarrollo de un organismo eucariótico se necesitan moléculas de ADN muy largas, lo que plantea un problema de empaquetamiento en un volumen pequeño, como lo es el núcleo celular.
En las células eucarióticas el ADN, incluso cuando se está duplicando, siempre se encuentra asociado a proteínas de carácter básico llamadas histonas, capaces de neutralizar las cargas ácidas del ADN. Al complejo de ADN y proteínas asociadas se le denomina cromatina.
Hay diferentes niveles de empaquetamiento de la cromatina a lo largo de las distintas etapas de la vida celular.
Las histonas se asocian con el ADN en un primer nivel de empaquetamiento para formar una fibra de 11 nm (110 A) llamada collar de perlas.
Si la fibra de 11 nm se digiere con una nucleasa que degrada el ADN no protegido, se liberan unas partículas: los nucleosomas.
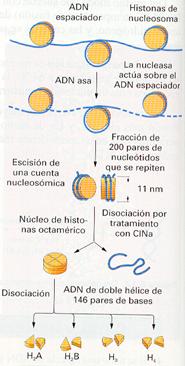
Estructura del collar de perlas y del nucleosoma.
Cada nucleosoma está formado por un núcleo con ocho moléculas de histona: dos de H2A, dos de H2B, dos de H3 y dos de H4. La cadena de ADN rodea el octámero dándole una vuelta y tres cuartos, ocupando 146 pares de nucleótidos.
En la célula en interfase generalmente la cromatina se vuelve a compactar mediante la histona H1 para formar la fibra de 30 nm. La fibra de 30 nm se empaqueta, a su vez, por otras proteínas en una serie de dominios de diferente grado de plegamiento.
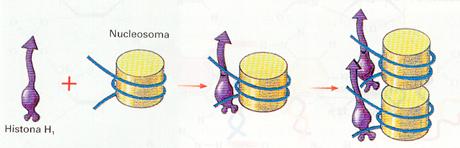
Estructura de la fibra de cromatina: Asociación del nucleosoma con la histona H, para constituir la fibra de 30 nm.
Hay dos tipos de cromatina funcional: la cromatina "activa" o eucromatina, que corresponde a las regiones activas con transcripción del ADN con una conformación relativamente relajada, y la cromatina más condensada o heterocromatina, que está inactiva.
En los momentos previos a la división celular, el material nuclear sufre otros empaquetarnientos sucesivos que aún no se conocen muy bien hasta alcanzar la estructura de los cromosomas metafásicos, en los que el nivel de empaquetamiento es máximo y llega a una 50.000 parte de la longitud inicial.
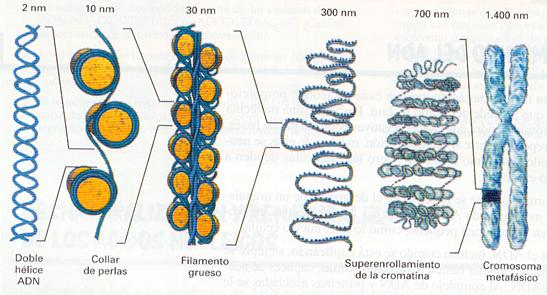
Empaquetamientos sucesivos.
Cada cromosoma se comporta como dos madejas de ADN, una por cada cromátida, de modo que el reparto equitativo a las células hijas se realice sin problemas.
ARN
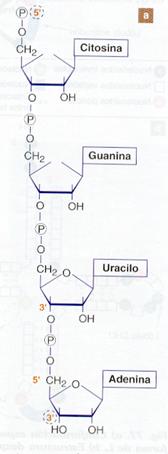
Está formado por cadenas muy largas de nucleótidos unidos entre sí por enlaces fosfodiéster entre las posiciones 3' y 5' de los nucleótidos consecutivos.
El azúcar es la ribosa, y las bases nitrogenadas son la adenina, la guanina, la citosina y el uracilo.
La molécula se repite monótonamente, y la única variable es la posición de las distintas bases nitrogenadas en la molécula. La cadena tiene polaridad, es decir, los dos extremos de la molécula son diferentes: uno es el extremo 5'fosfato, y el otro es el extremo 3'hidroxilo. Por convenio, siempre que no se especifique la polaridad, las cadenas se escriben de 5' a 3'.
- El orden de los nucleótidos en la molécula de ARN se llama secuencia y constituye la estructura primaria del ARN. La secuencia de un ARN determina un mensaje genético.
- Las bases nitrogenadas pueden formar puentes de hidrógeno entre sí; preferentemente, adenina con uracilo, y citosina con timina. La molécula se estabiliza cuando ha formado la mayor cantidad de puentes de hidrógeno. Esto le proporciona una estructura determinada: la estructura secundaria.
- Esta estructura puede plegarse a su vez en el espacio obteniendo una configuración específica estable. Ésta es la estructura terciaria de la molécula.
|
|
a) Fragmento de ARN. b) El ARN es una hebra sencilla que puede presentar cortas zonas de apareamiento de bases |
Existen cuatro tipos de ARN, relacionados de una forma u otra con la expresión de la información genética contenida en el ADN. Estos tipos son el ARN mensajero, ARN ribosómico, ARN de transferencia y ARN nucleolar.
ARN mensajero (ARNm)
ElARNm es una copia de una parte del ADN (la que corresponde a cada gen o grupo de genes que vaya a expresarse) que será utilizada por los ribosomas como información para poder unir los aminoácidos en el orden adecuado y constituir una proteína concreta.
Las cadenas de ARN mensajero tienen una vida muy corta, ya que si no fueran destruidas (mediante enzimas ribonucleasas) la síntesis proteica se prolongaría indefinidamente y se originaría una superproducción de proteínas. Así, cuando se necesita sintetizar una proteína concreta, se fabrica de nuevo el ARN mensajero correspondiente.
ElARNm constituye, aproximadamente, entre el 3 y el 5 % del total del ARN celular.
ARN transferente (ARNt)
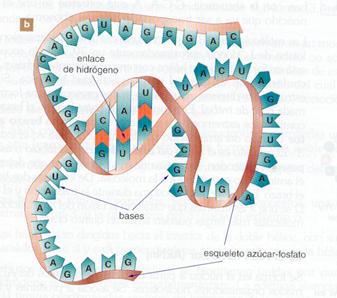
El ARNt se encarga de transportar los aminoácidos presentes en el citoplasma celular hasta los ribosomas, donde se unirán para constituir las proteínas. Cada molécula de ARNt transporta un aminoácido específico. Estas diferencias son debidas, fundamentalmente, a una secuencia de tres bases nitrogenadas, denominada anticodón, que varía entre los distintos ARNt.
Los ARNt están formados por cadenas cortas (entre 70 y 90 nucleótidos) que contienen un 10% de bases nitrogenadas diferentes a las cuatro mayoritarias, como ya se ha indicado.
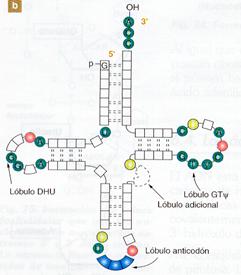
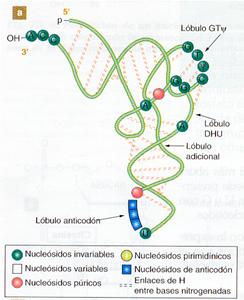
Las moléculas de ARNt poseen una estructura secundaria muy característica, en la que existen tramos de doble cadena, por emparejamiento intracatenario. Estos tramos se denominan brazos y hay cuatro en cada molécula, aunque también logra aparecer un quinto brazo más corto que los otros. En los extremos de tres de los brazos existen unas zonas sin emparejar que componen los denominados bucles.
El extremo 3' de la cadena tiene siempre la secuencia de bases CCA. A este nucleótido terminal de adenina se une el aminoácido que va a ser transportado. En el extremo 5' siempre existe un nucleótido con guanina.
La estructura extendida del ARNt tiene forma de hoja de trébol, aunque en la realidad los brazos se disponen plegados, constituyendo una estructura acodada que recibe el nombre de «estructura en boomerang».
El ARNt constituye, aproximadamente, el 10 % del ARN celular total.
|
|
|
|
a) Conformación espacial de ARNt. |
b) Estructura desplegada en forma de “ hoja de trébol" |
ARN ribosómico (ARNr)
El ARNr forma parte de los ribosomas y participa, por tanto, en el proceso de unión de los aminoácidos para sintetizar las proteínas. No constituye una molécula específica de cada organismo, pues, a diferencia del ARNm, no contiene información sobre la clase de proteína que se va a sintetizar.
Existen varias cadenas distintas de ARNr, que se diferencian por su coeficiente de sedimentación.
Las células procariotas y eucariotas tienen cadenas de ARNr ligeramente distintos.
El ARNr es el tipo más abundante, ya que corresponde al 80-85% del ARN celular total.
El ARN nucleolar (ARNn)
Se forma en el núcleo a partir de ciertos segmentos del ADN llamados organizadores nucleolares. Se asocia a proteínas y forma el nucléolo. Después se fragmenta y da las subunidades de los ribosomas, que salen por los poros nucleares hacia el citoplasma.
LOCALIZACIÓN DE LOS ÁCIDOS NUCLEICOS.
En los eucariotas, el ADN se encuentra localizado en el núcleo formando largas moléculas lineales asociadas a proteínas básicas. Aunque la mayor parte del ADN de las células eucariotas está confinada en el núcleo, en las mitocondrias y en los cloroplastos también hay ADN.
En los eucariotas el ARN se localiza tanto en el núcleo como en el citoplasma. Todos los ARN (ARN mensajero, ARN ribosómico, ARN de transferencia y ARN nucleolar) se sintetizan en el núcleo utilizando como molde una de las dos cadenas de polinucleótidos del ADN.
- El ARN mensajero, una vez sintetizado, atraviesa la membrana nuclear y pasa al citoplasma.
- El ARN ribosómico, denominado también ARN estructural, se asocia a un conjunto de proteínas básicas y forman los ribosomas.
- El ARN de transferencia se localiza en el citoplasma, desde donde transporta los aminoácidos hasta el ribosoma para que se unan y formen las proteínas.
- El ARN nucleolar se encuentra asociado a diferentes proteínas formando el nucléolo. Se origina en el núcleo a partir de diferentes segmentos del ADN denominados organizadores nucleolares. Una vez formado, se fragmenta y da origen a los diferentes tipos de ARN ribosómico.
En las procariotas, al carecer de núcleo todos los ácidos nucleicos están en el citoplasma o formando parte de los ribosomas; también se puede encontrar ADN en unas formaciones llamadas plásmidos...
En los virus tanto el ADN como el ARN pueden formar parte de su material genético, pero no juntos.
FUNCIONES DE LOS ACIDOS NUCLEICOS.
El ADN es el portador del mensaje genético, que ha de pasar invariable de una generación celular a otra. Para eso, ha de duplicarse transmitiendo el mismo mensaje en las dos copias. A este proceso se le llama replicación.
El mensaje genético contenido en el ADN es la especificación de las proteínas que ha de sintetizar la célula. Desde el mensaje del ADN hasta las moléculas de proteínas se pasa por dos procesos:
· transcripción, que consiste en que el mensaje de un fragmento de ADN es copiado (transcrito) en una molécula de ARN. Es decir, consiste en la síntesis de una molécula de RNA que contenga el mismo mensaje que el correspondiente fragmento de ADN.
· traducción, que consiste en la síntesis de la proteína significada por el mensaje del ADN, siguiéndose las instrucciones del ARN.
Para que se lleve a cabo la traducción, han de intervenir tres tipos diferentes de ARN, que son:
- el ARN mensajero (ARNm), que dicta las órdenes de colocación de los aminoácidos, pues reproduce el mensaje genético de un fragmento de ADN.
- el ARN transferente (ARNt), que transporta los aminoácidos hasta las moléculas de mRNA.
- el ARN ribosómico (ARNr), tiene función estructural, pues, forma parte de los ribosomas, que son los orgánulos celulares donde se realiza el acoplamiento entre el ARNm y el ARNt, y la unión de los aminoácidos para formar las proteínas, y por tanto constituyen el lugar donde se realiza la traducción.
El RNA nucleolar (nRNA), se encuentra asociado a proteínas formando el nucleolo.
Algunos virus carecen de ADN y, por ello, contienen su información biológica en forma de ARN.
*Observar que en esta pregunta la forma de escribir la abreviatura de los ácidos nucleicos es distinta, pero válida, aunque menos usada.
Aparte de las estructuras que hemos visto de los ácidos nucleicos existen OTRAS ESTRUCTURAS, pudiendo resumir de la siguiente manera todas las estructuras posibles: