



Dinámica del ecosistema
5.1.MECANISMOS DE AUTORREGULACIÓN
El tamaño de una población viene condicionado por los factores limitantes y por estrategias reproductoras propias de cada especie.
Factores limitantes. Son aquellos que condicionan el aumento de población, afectando a las tasas de mortalidad o natalidad o bien a ambas. Distinguimos dos tipos de factores limitantes:
Factores dependientes de la densidad. Se originan por un aumento excesivo de la población, lo que produce un aumento de la competencia por el alimento, el espacio, reproducción, enfermedades... Todo conduce a disminuir el tamaño de la población.
Factores independientes de la densidad. Afectan a las tasas de natalidad y mortalidad sin tener en cuenta el tamaño de la población. Suelen ser factores abióticos como una inundación, una erupción volcánica, una sequía…
5.1.1 FACTORES ABIÓTICOS LIMITANTES: ESPECIES "ESTENO" Y "EURI". LÍMITES DE TOLERANACIA
Cada especie de ser vivo necesita ciertas condiciones fisicoquímicas para vivir. Tales condiciones pueden ser muy variadas dependiendo de las características del organismo y pueden variar enormemente de un organismo a otro. Uno solo de los factores ambientales puede ser suficiente para que un ser vivo no pueda vivir en un área determinada, de acuerdo con la ley del Mínimo de Liebig (la distribución de una especie está controlada por aquel factor ambiental para el cual el organismo tiene la mínima capacidad de adaptación). Es precisamente el factor o requerimiento más escaso (limitante) el que, con su presencia o ausencia, regula la supervivencia de los organismos y por tanto el tamaño de la población.
La presencia o la actividad vital de los organismos está limitada por un cierto intervalo de valores para cada factor ambiental. Cuando se representa gráficamente el número de individuos de una población frente a intensidades variables del factor considerado, se obtiene una curva en forma de campana que se llama curva de tolerancia.
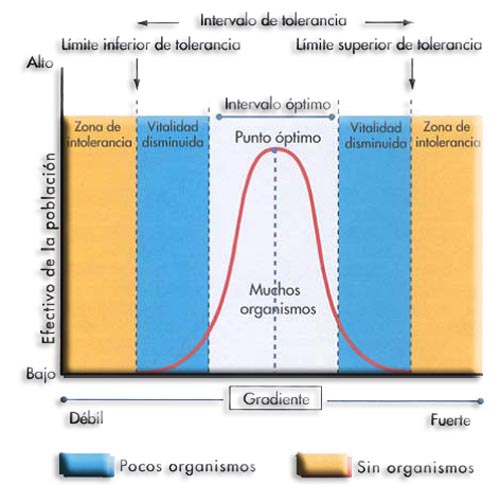
La parte central de la curva representa el intervalo de valores óptimos para el desarrollo de los organismos, y coincide con el mayor tamaño de la población. El punto de inflexión se corresponde con el punto óptimo. Los extremos de la curva se corresponden con los límites de tolerancia mínimo y máximo, si se sobrepasan, los organismos mueren.
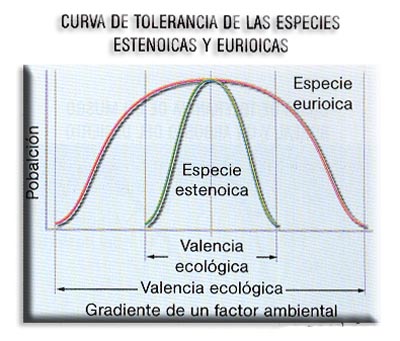
Los seres vivos que habitan en un medio están continuamente sometidos a una serie de factores ambientales. Denominamos valencia ecológica al intervalo de tolerancia de una especie respecto de un factor del medio (luz, Tª, humedad, pH...) que actúa como factor limitante; o sea, es la capacidad de un organismo para poblar medios diferentes.
Desde el punto de vista de la amplitud de la valencia ecológica, podemos considerar dos tipos de especies diferentes:
a) Especies estenoicas. Son aquellas que presentan un estrecho margen de tolerancia para un factor determinado. Así hablamos de organismos hidrófilos, que necesitan un alto grado de humedad para vivir (anfibios adultos, musgos…) y xerófilos (cactus), adaptados a vivir sin agua. En un caso la escasez y en otro la abundancia de agua son nocivos para estos organismos.
Se habla de organismos estenotermos, estenohalinos o estenohigros, para indicar su estrecha tolerancia a la temperatura, la salinidad o a la humedad, respectivamente. Son especies muy exigentes que necesitan para vivir unos límites muy estrechos, por lo que tienen una valencia ecológica pequeña.
b) Especies eurioicas. Presentan un amplio margen de tolerancia, esto es, se acomodan a condiciones muy variables. La mayoría de las especies de nuestras regiones tienen moderadas necesidades de agua, soportando bien la alternancia de estaciones húmedas y secas o con altas y bajas temperaturas. Todas las especies que habitan en los estuarios y marismas son especies eurihalinas pues están adaptadas a soportar grandes y rápidos cambios de salinidad.
Se habla de euritermos, eurihalinos o eurihigros para indicar su amplia tolerancia a la temperatura, a la salinidad o a la humedad, respectivamente. Su valencia ecológica es muy grande.
5.1.2 ESPECIES ESTRATEGAS DE LA "K" Y DE LA "R"
Hemos visto que las especies eurioicas tienen valencias ecológicas de gran amplitud, sin embargo, las estenoicas tienen valencias muy estrechas. Por lo tanto, las especies menos tolerantes suelen responder de manera más eficaz cuando las condiciones del medio le son propicias, siendo más especialistas o "k" estrategas; por el contrario, las especies eurioicas son generalistas o "r" estrategas.
Estrategas de la "r" (reproducción). Son aquellos organismos dotados de un alto potencial biótico con unas tasas de natalidad y de mortalidad elevadas. Su estrategia consiste en asegurar la descendencia con
elevadas tasas de reproducción. Son especies oportunistas que colonizan rápidamente hábitat con unas condiciones que cambian en poco tiempo (charcas, zonas semidesérticas...). Soportan mal la competencia, a la que, frecuentemente, responden emigrando.
Invierten gran cantidad de materia y energía en la producción de ingentes cantidades de huevos o semillas capaces de asegurar la reproducción a pesar de que la gran mayoría de ellos morirá tempranamente. Los supervivientes se reproducen rápidamente, repitiendo el ciclo.
Muchos organismos pequeños como bacterias, algas y hongos se incluyen en este grupo, que también comprende a numerosos insectos y a pequeños vertebrados como los ratones, topillos...
Están mínimamente adaptados a las variaciones del medio, por lo que están expuestos a tasas de mortalidad catastróficas. Su desarrollo es rápido y alcanzan muy pronto la madurez sexual por lo que son poco longevos (menos de 1 año) y suelen tener muchas crías que se cuidan solas.
Estrategas de la "k" (capacidad de carga del ambiente). Son organismos especialistas que prefieren hábitats estables con condiciones ambientales constantes. Las 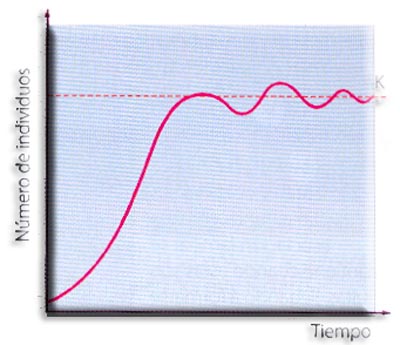 características biológicas de la especie determinan la regulación de la población más que las condiciones del medio. Frecuentemente alcanzan su máxima capacidad de carga, regulándose las poblaciones mediante competencia, migraciones...
características biológicas de la especie determinan la regulación de la población más que las condiciones del medio. Frecuentemente alcanzan su máxima capacidad de carga, regulándose las poblaciones mediante competencia, migraciones...
Tienen poca descendencia y solo tras largos periodos (más de 1 año) alcanzan talla y peso considerable. Su tiempo de vida es en general largo, y los progenitores suelen dedicar grandes cuidados a sus crías.
Son k-estrategas la mayoría de las aves y mamíferos y los grandes árboles. Si no es posible la coexistencia, los estrategas de la “k” desplazan a los de la “r”.
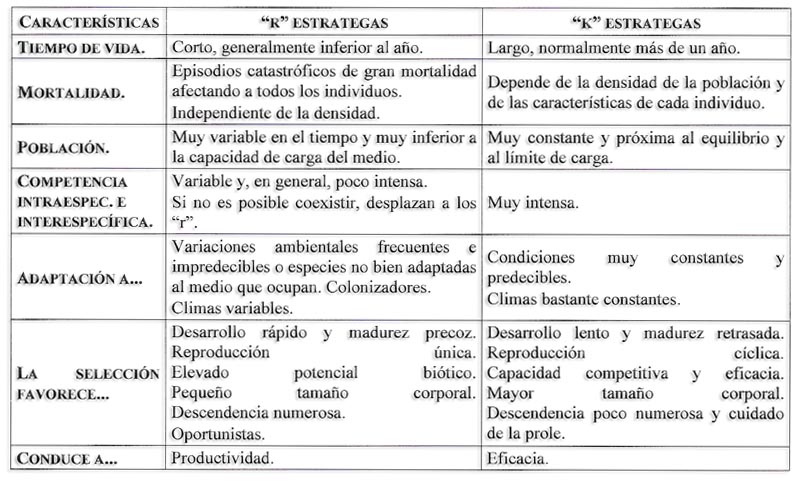
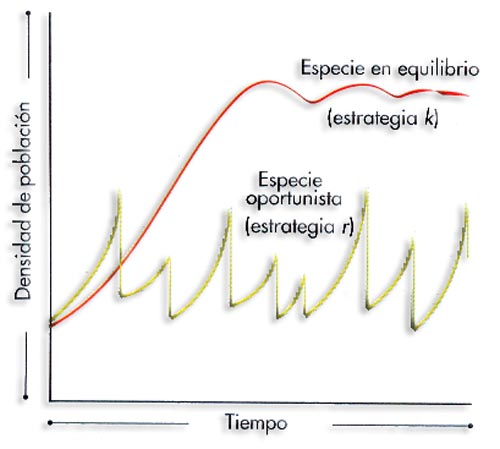
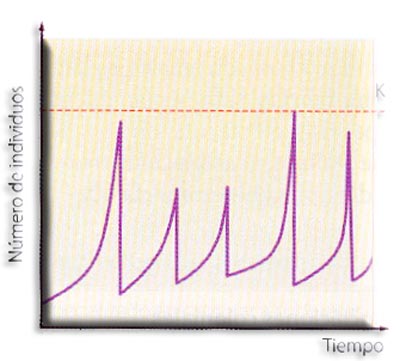
Curva de crecimiento de la población de una especie en equilibrio frente a la de una especie oportunista, sujeta a una mortalidad catastrófica irregular
5.1.3 DINÁMICA DE POBLACIONES: CRECIMIENTO DE POBLACIONES
Hay una serie de parámetros que determinan como se desenvuelve una población, entre ellos destacan: el tamaño de la población (número de individuos), la distribución de edades, la densidad y distribución espacial y las estrategias de regulación del tamaño de la población.
5.1.3.1 TAMAÑO DE POBLACIÓN
El tamaño de una población viene determinado por el número de individuos de la misma. Hay una serie de factores que tienden a favorecer el aumento o la disminución de la población, entre ellos se encuentran:
Tasa de natalidad. Es el número de individuos nacidos por unidad de tiempo.
Tasa de inmigración. Es el número de individuos que se suman a una población procedente de otras poblaciones.
Tasa de mortalidad. Número de individuos que mueren en una población.
Tasa de emigración. Número de individuos que abandonan una población para dirigirse a otros ecosistemas.
5.1.3.2 CRECIMIENTO DE LA POBLACIÓN
Existen dos fuerzas que actúan sobre el crecimiento de la población: el potencial biótico (r) (capacidad de reproducirse a un cierto ritmo), característica inherente a cada población, y la resistencia ambiental, constituida por todos los factores físicos y biológicos que frenan el crecimiento de la población.
Considerando todos estos parámetros se puede calcular el crecimiento de una población.
Crecimiento exponencial o en "J". Si las hembras de una especie se cruzan con los machos y si todos los descendientes sobreviven y alcanzan la madurez, la población crece muy rápidamente. Dicho de otro modo, si la tasa de natalidad es máxima y la de mortalidad mínima, no hay límites que pongan freno a la población, por lo que la población crecerá de forma desmesurada siguiendo una ecuación exponencial. Su gráfica será una curva exponencial o en "J".
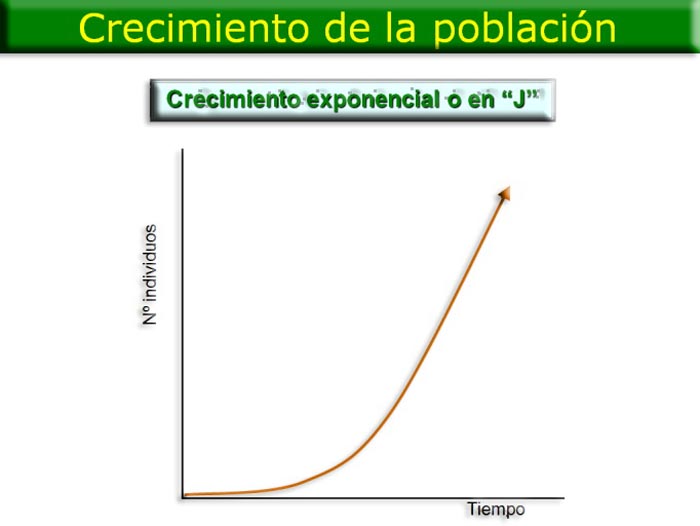
El tipo de crecimiento exponencial es propio solo de microorganismos que crecen en condiciones óptimas en el laboratorio. En la naturaleza, cualquier población no puede creer indefinidamente ya que los recursos vitales disponibles (comida, agua, espacio...) se agotarían muy rápidamente, y las relaciones de competencia que se establecen entre sus miembros harían que la población disminuyese rápidamente. El conjunto de todas estas interacciones de los organismos con el medio y entre ellos, que determinan el crecimiento de la población, se denomina resistencia ambiental.
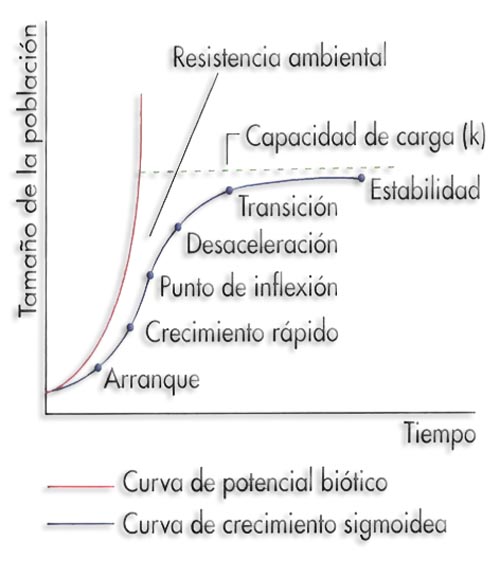
Representación comparativa del crecimiento de una población según su potencial biótico y del crecimiento real
Crecimiento sigmoideo o logístico. Para comprender la relación entre el potencial biótico y la resistencia ambiental, vamos a estudiar el crecimiento de las levaduras en el laboratorio.
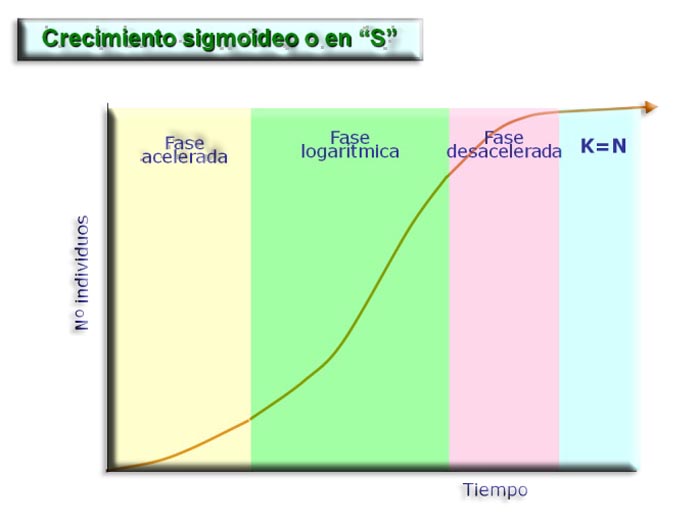
Inicialmente el crecimiento es muy lento (fase de latencia), le sigue una fase de crecimiento rápido (fase exponencial) que se ralentiza nuevamente cuando la población alcanza cierto tamaño (fase estacionaria). El crecimiento rápido tiene lugar durante las primeras 8 horas y después disminuye hasta hacerse 0, lo que indica que la población está en equilibrio con el medio, es decir, se ha alcanzado el tamaño de la población que el medio puede sostener. Este número máximo de individuos de una población que puede soportar el medio se denomina capacidad de carga (K). Una vez alcanzada esta fase de equilibrio, la población se estabiliza compensándose las tasas de natalidad y mortalidad. De esta manera, el número de individuos fluctúa en torno a la capacidad de carga. K = N (crecimiento cero).
La mayoría de animales y plantas sigue una curva de crecimiento sigmoideo en "S".
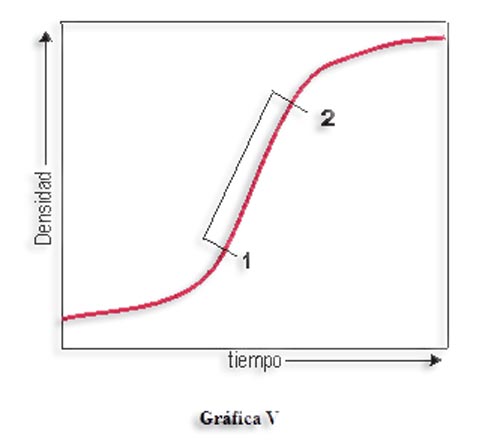
La explotación de los ecosistemas por el hombre (agricultura, ganadería o pesca), consiste en extraer biomasa manteniendo el ecosistema inmaduro. Para ello hay que evitar que progrese la sucesión y que el consumo respiratorio suponga una menor producción neta. Desde el punto de vista de la demografía se trataría de mantener la población en ese segmento 1-2 de crecimiento exponencial, evitando que el aumento de la densidad haga decrecer la producción. Pero la sobreexplotación significa extraer más deprisa de lo que puede crecer la población, por lo que se reducirá su densidad a un nivel inferior al de producción óptima (antes de alcanzar el punto 1). El buscar el máximo beneficio en el menor plazo posible puede conducir a reducir los niveles de la población objeto de explotación por debajo de ese límite que permita la recuperación de la misma.
5.1.4 RELACIONES INTERESPECÍFICAS
Son relaciones existentes entre dos especies o poblaciones diferentes de una misma comunidad. En estas relaciones, una especie puede salir beneficiada, perjudicada o permanecer indemne.
a) Depredación (+ -). Es una relación en la que un organismo vivo, la presa, es matado y consumido total o parcialmente por otro que se beneficia, el depredador. E1 modelo depredador-presa es estabilizador ya que se basa en la existencia de un bucle de realimentación negativo.
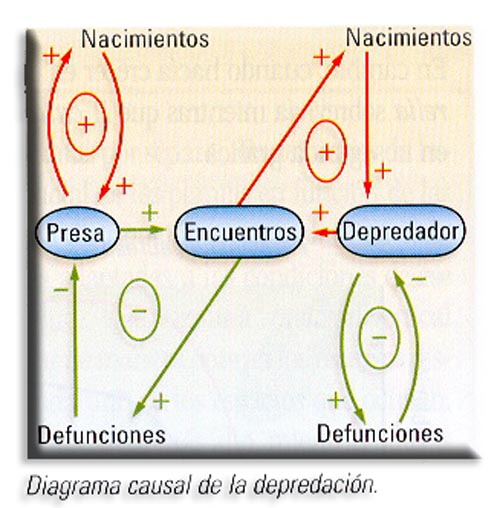
Los depredadores pueden llegar a exterminar a las poblaciones de la presa, al menos en potencia; sin embargo, esta es una consecuencia improbable ya que por lo general las poblaciones del depredador y de la presa siguen un modelo oscilatorio pero ligeramente desfasado.
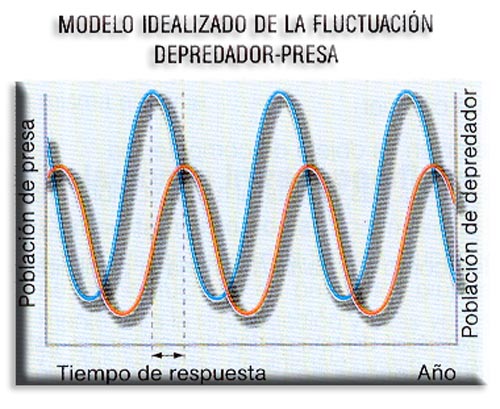
Estas fluctuaciones solo son patentes cuando el depredador consume presas de una sola especie. Los ecosistemas ofrecen refugios apropiados a las presas, lo que dificulta su captura por los depredadores. Es este un factor decisivo para la interacción entre ambas poblaciones que evita así la extinción de cualquiera de ellas.
La selección natural actúa aumentando la eficacia del depredador para capturar el alimento y, al mismo tiempo, aumenta la eficacia de la presa para escapar del depredador.
Desde el punto de vista funcional podemos considerar dos tipos de depredadores: carnívoros (caracterizados por dar muerte rápidamente a sus presas) y ramoneadores (organismos herbívoros).
 Las plantas carnívoras. Son vegetales que se nutren parcialmente de animales, en especial de insectos, que capturan ellas mismas. Casi todas crecen en turberas y humedales, donde el suelo es ácido y pobre en nitrógeno asimilable. La falta de este elemento imprescindible les impide elaborar las proteínas que precisan. Por ello han tenido que desarrollar mecanismos para aprovechar la escasa fuente de nitrógeno de estas zonas: las proteínas de los animales, principalmente insectos, las asimilan por medio de enzimas digestivas especiales y de bacterias que descomponen sus cuerpos. Los minerales disueltos son absorbidos por la planta que recibe así raciones extra de nitrógeno y fósforo. Al mismo tiempo, las hojas verdes de estas plantas fabrican hidratos de carbono mediante la fotosíntesis (autótrofos).
Las plantas carnívoras. Son vegetales que se nutren parcialmente de animales, en especial de insectos, que capturan ellas mismas. Casi todas crecen en turberas y humedales, donde el suelo es ácido y pobre en nitrógeno asimilable. La falta de este elemento imprescindible les impide elaborar las proteínas que precisan. Por ello han tenido que desarrollar mecanismos para aprovechar la escasa fuente de nitrógeno de estas zonas: las proteínas de los animales, principalmente insectos, las asimilan por medio de enzimas digestivas especiales y de bacterias que descomponen sus cuerpos. Los minerales disueltos son absorbidos por la planta que recibe así raciones extra de nitrógeno y fósforo. Al mismo tiempo, las hojas verdes de estas plantas fabrican hidratos de carbono mediante la fotosíntesis (autótrofos).
Para ser carnívora, una planta debe tener las siguientes características:
- Atraer a la presa: (olor, color, néctar...).
- Tener adaptaciones especificas para atrapar la presa: (trampa de fosa bisagra, de papel matamoscas....).
- Tener adaptaciones para utilizar la presa (enzimas digestivas, hongos simbióticos, bacterias...).
b) Parasitismo (+ -). Es una relación binaria en la que un individuo, el parásito, vive a expensas de otro que es el hospedador, que resulta perjudicado.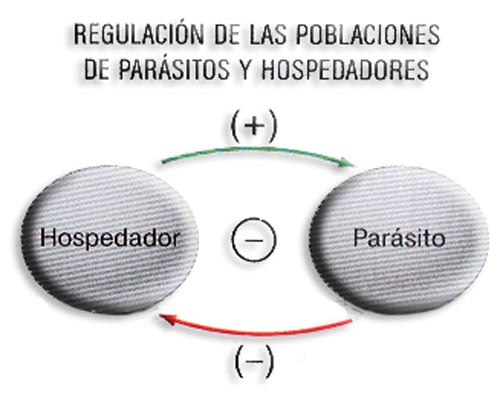
Este modelo es similar al D/P salvo que, en la mayoría de las ocasiones, al parásito no le interesa matar al hospedador; o sea, el depredador se mantiene del "capital" mientras que el parásito lo hace de los "intereses", por lo que no le interesa acabar con la victima pues supondría su fin.
Puede haber dos clases de parasitismo:
Endoparasitismo. El parásito vive dentro del organismo hospedante (duela del hígado, tenia...).
Ectoparasitismo. El parásito es externo (pulga, piojo, mosquito...). Entre los vegetales podemos citar algunos fitoparásitos como el muérdago que parasita las ramas del pino o el jopo que parásita las raíces del trébol.
El piojo de mar (Anilocra) es un crustáceo que se adhiere a los peces haciendo uso de las afiladas garras de sus patas. Una vez sujeto, se alimentará de la sangre de su nuevo huésped. Algunas aves como el cuco depositan los huevos en el nido de otra especie. Los huevos del hospedador son eliminados por los pollos del cuco con el fin de que los huevos del parásito reciban los cuidados que necesitan para desarrollarse, suplantando a los huevos del hospedador. El parásito ha evolucionado llegando a mimetizarse para que la especie parasitada no rechace los huevos extraños.
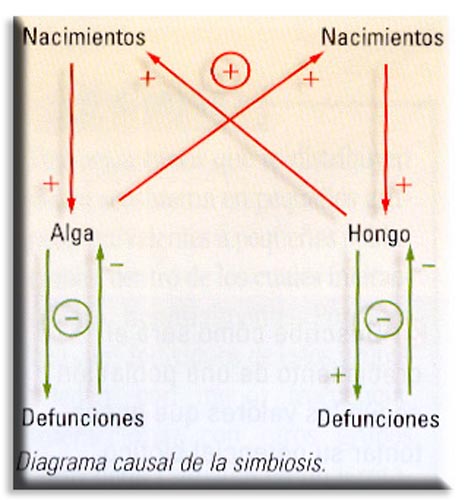 c) Simbiosis (+ +). Se da cuando dos organismos se asocian para vivir en comunidad obteniendo un beneficio mutuo. La asociación es tan íntima y permanente que forma un todo orgánico. Son casos de simbiosis la asociación entre determinadas algas y hongos para formar líquenes. El alga recibe un lugar protegido con la humedad necesaria y el hongo recibe productos orgánicos para su nutrición. En la agricultura es muy normal aprovechar esta ventaja de la simbiosis que se da en las plantas leguminosas. Estas albergan en sus raíces bacterias nitrificantes del género Rhizobium que fijan el nitrógeno atmosférico y se lo ceden a la planta, permitiendo rotar los cultivos y aprovechar el suelo nitrogenado. La leguminosa aporta a la bacteria un lugar protegido y nutrientes orgánicos. Otro ejemplo es el de los insectos xilófagos como las termitas y los protozoos del intestino que digieren la celulosa.
c) Simbiosis (+ +). Se da cuando dos organismos se asocian para vivir en comunidad obteniendo un beneficio mutuo. La asociación es tan íntima y permanente que forma un todo orgánico. Son casos de simbiosis la asociación entre determinadas algas y hongos para formar líquenes. El alga recibe un lugar protegido con la humedad necesaria y el hongo recibe productos orgánicos para su nutrición. En la agricultura es muy normal aprovechar esta ventaja de la simbiosis que se da en las plantas leguminosas. Estas albergan en sus raíces bacterias nitrificantes del género Rhizobium que fijan el nitrógeno atmosférico y se lo ceden a la planta, permitiendo rotar los cultivos y aprovechar el suelo nitrogenado. La leguminosa aporta a la bacteria un lugar protegido y nutrientes orgánicos. Otro ejemplo es el de los insectos xilófagos como las termitas y los protozoos del intestino que digieren la celulosa.
d) Competencia (- -). Dos poblaciones compiten cuando utilizan el mismo recurso que puede llegar a escasear y, por tanto, limitar el desarrollo de ambas poblaciones. Si los requerimientos son muy similares, mayor será la competencia, y únicamente sobrevivirá la población de una de las especies, mientras que la otra queda excluida. Se trata de un principio de exclusión competitiva.
- Competencia intraespecífica. Este tipo de relación se da entre individuos de la misma especie (chopos que crecen muy juntos cuyas ramas compiten por la luz y sus raíces por el agua y las sales minerales). La competencia intraespecífica es más fuerte ya que consiste en la lucha por unos recursos idénticos. En ella se cumple el "principio de exclusión competitiva" que dice que "dos especies no pueden ocupar simultáneamente y de forma permanente un mismo nicho ecológico".
- Competencia interespecífica. Se da entre especies distintas (ovejas y cabras que conviven en un mismo territorio).
Para evitar la competencia y explotar completamente los recursos, las especies diversifican sus comportamientos de diferentes formas:
- Practicando formas distintas de alimentación. Así el gamo pasta herbáceas en el suelo y el ciervo ramonea hojas de los árboles.
- Compartiendo un mismo alimento de manera escalonada. Jirafas, impalas y gacelas conviven en las sabanas alimentándose de las acacias, pero a distinta altura.
- Criando en diferentes épocas del año. El petirrojo cría en abril, el colirrojo real en mayo, y el papamoscas en junio, pero los tres son insectívoros.
- Migrando. Así se explota el mismo ecosistema pero por especies distintas y en diferentes épocas del año.
Para evitar la competencia y explotar completamente los recursos, las especies diversifican sus comportamientos de diferentes formas:
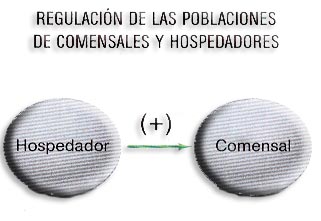 e) Comensalismo (+ =). Es una relación en la que una especie se aprovecha del sobrante de la comida u otros productos de otra especie que denominamos patrón. Ninguna de las dos especies resulta perjudicada, al contrario, una obtiene un beneficio y la otra, el patrón (especie controlante), ni beneficio ni perjuicio.
e) Comensalismo (+ =). Es una relación en la que una especie se aprovecha del sobrante de la comida u otros productos de otra especie que denominamos patrón. Ninguna de las dos especies resulta perjudicada, al contrario, una obtiene un beneficio y la otra, el patrón (especie controlante), ni beneficio ni perjuicio.
Por ejemplo: el cangrejo y la actinia, los animales carroñeros (se asocian a los carnívoros para apoderarse de sus despojos), animales arborícolas y aves (viven sobre los árboles utilizándolos como soporte), u organismos que viven en los nidos de las aves aprovechando los restos de alimento. En otros casos, el comensal se aprovecha de las mudas, descamaciones o excrementos del patrón.
f) Mutualismo (+ +). Es una relación en la que ambos organismos resultan beneficiados. A diferencia de la simbiosis, los dos organismos pueden vivir de forma independiente. En la mayoría de los casos es una asociación trófica. Son ejemplos de mutualismo las relaciones entre las gaviotas y el hombre. Estas se alimentan de los restos de actividades pesqueras realizando una limpieza beneficiosa para el hombre.
Otros ejemplos son la agricultura (hombre-plantas), la ganadería (hombre-animales), los insectos polinizadores y las plantas, los pulgones y las hormigas, las garcillas bueyeras (acompañan al ganado al que desparasitan), las especies limpiadoras (generalmente gambas y pequeños peces de la familia de los Lábridos) que se alimentan de los parásitos que se hospedan en la piel de otros peces. De este modo, cuando un pez llega a una zona de limpieza suele adoptar una postura que invita a los pequeños limpiadores a comenzar su tarea.
g) Foresia (+ =). En ella, un organismo pequeño invade a otro de mayor tamaño para utilizarlo como medio de transporte o diseminación sin ocasionarle ningún daño. Como ejemplo tenemos las rémoras (peces provistos de una ventosa cefálica con la que se sujetan a especies mayores como tiburones), que son transportadas de un lugar a otro; los cirrípedos que se fijan sobre la piel de los cetáceos, los pájaros que transportan en su intestino semillas de plantas...
h) Tanatocresis (+ =). Es una relación en la que una especie obtiene un beneficio pero este no es trófico. Consiste en la utilización de cadáveres, piezas esqueléticas, excrementos y otros productos de una especie por individuos vivos de otra especie pero no como alimento. Con los restos pueden construirse refugios (el cangrejo ermitaño se aloja en conchas abandonadas de moluscos, las larvas de las frigáneas construyen tubos con restos de ramas para vivir en su interior) o emplearse de modos diferentes como es el caso del pez payaso que vive entre los tentáculos cargados de células urticantes de muchos celentéreos.
i) Epibiosis (+ =). Es una relación, permanente o no, en la que una especie se sirve de otras como sustrato de fijación, generalmente sin perjuicio para ella o con cierto beneficio (camuflaje). Así muchos cangrejos recubren su caparazón de algas, al igual que ocurre con las conchas de los mejillones, ostras...
5.2 SUCESIÓN Y REGRESIÓN DE LOS ECOSISTEMAS
Como hemos visto, las poblaciones, las comunidades y las condiciones fisicoquímicas del biotopo varían a lo largo del tiempo. En consecuencia, los ecosistemas cambian pues no son entidades que permanecen estáticas; además, son capaces de mantener e incrementar continuamente su organización, reajustándose y adaptándose a cualquier tipo de variación, utilizando un flujo continuo de materia y energía.
El final de este camino es un estado teórico de máxima madurez y estabilidad dinámica que se denomina clímax. El proceso que siguen todos los ecosistemas tratando de alcanzar su clímax recibe el nombre de sucesión ecológica. Se puede definir como "un proceso dinámico resultante de las interacciones entre los factores bióticos y abióticos en el tiempo, que da lugar a la formación de un ecosistema complejo y estable".
Como consecuencia de dicho proceso, en los ecosistemas hay sustituciones de sus biocenosis a lo largo del tiempo, sucediéndose cronológicamente unas a otras y proporcionando cada una de ellas condiciones favorables para la siguiente. La sucesión ecológica es un proceso lento y gradual en el que las comunidades inestables sufren modificaciones, tanto en su composición como en el tamaño de sus poblaciones, buscando el equilibrio con los factores abióticos del ecosistema.
Cuando este equilibrio se logra en la etapa final de la sucesión (clímax), la comunidad tendera a mantenerse, no será sustituida por ninguna otra mientras no cambien las condiciones fisicoquímicas y climáticas. Así, en las zonas con climas más constantes de la Tierra encontramos los ecosistemas más maduros y complejos: la selva tropical y los arrecifes coralinos.
Si ocurriese un cambio, el clímax se rompería y el ecosistema iniciaría otra sucesión hasta alcanzar un nuevo clímax. Este proceso de vuelta atrás se denomina regresión o disclimax, y puede ocurrir por causas naturales (cambio climático, inundaciones, erupciones volcánicas...) o antrópicas (deforestación, contaminación, introducción de especies nuevas en el ecosistema…).
5.2.1 TIPOS DE SUCESIONES
Las sucesiones pueden ser primarias y secundarias.
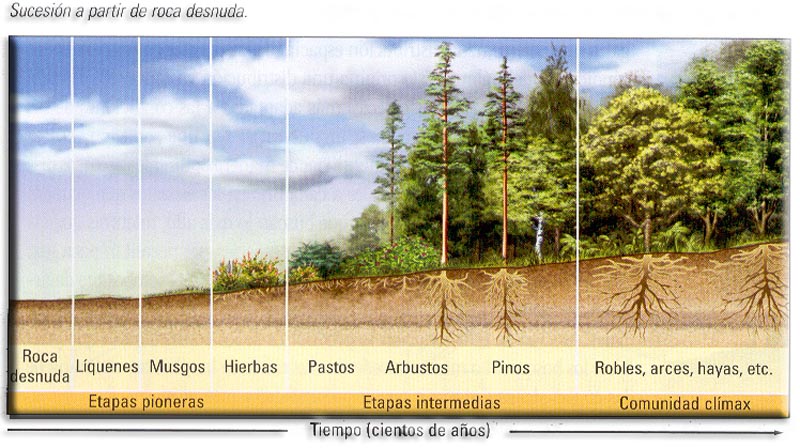
Sucesiones primarias. Se producen en territorios vírgenes que no han sido colonizados antes por ninguna comunidad. Es el caso de las dunas, coladas volcánicas, aluviones... Los primeros organismos que colonizan estos territorios son los líquenes y musgos, y a partir de ellos se empieza a formar suelo. Posteriormente entran bacterias y hongos y las primeras herbáceas con raíces superficiales.
Sucesiones secundarias. Ocurren en ecosistemas que han sufrido una regresión que ha interrumpido su camino hacia el clímax o en los que se ha roto dicho clímax pero todavía conservan el suelo y parte de la vegetación. Al cabo de un tiempo más o menos largo, si las condiciones ambientales son las mismas que las iniciales, el ecosistema vuelve a seguir la misma sucesión ecológica.
Así, por ejemplo, si después de cortar un bosque abandonamos el terreno, la vegetación puede volver a adueñarse de él. Las sucesiones secundarias no repiten necesariamente la misma serie que se dio en la primaria, ya que el medio puede haber sido totalmente modificado en mayor o menor grado. Así, en el caso de la tala del bosque, es posible que el suelo prácticamente no se altere. De todos modos, las primeras plantas que se desarrollan no suelen ser las del bosque, sino plantas herbáceas de crecimiento rápido que resisten la insolación directa. Posteriormente, a medida que se desarrollan árboles y arbustos, aparecerán también las plantas de sombra propias del bosque.
No todas las sucesiones exigen muchos años para su desarrollo. Sobre un tronco caído, por ejemplo, tiene lugar una rápida sucesión, y lo mismo ocurre sobre la carroña de un animal muerto. En cuanto a la vegetación terrestre, si dejamos aparte las sucesiones secundarias que hayan podido seguirse íntegramente, en general no es posible observar directamente toda la serie, especialmente por la lentitud de las etapas iniciales.
5.2.1.1 LOS CAMBIOS DE LAS SUCESIONES HACIAL EL CLÍMAX
A medida que transcurren, se puede apreciar una serie de cambios en los ecosistemas:
a) Aumento de la estabilidad. Las relaciones entre las especies que integran la biocenosis son muy fuertes, existiendo múltiples circuitos y realimentaciones que contribuyen a la estabilidad del sistema, amortiguándose las fluctuaciones.
b) Aumento de la biomasa. Se manifiesta sobre todo en los organismos con un metabolismo bajo. Por ejemplo, la "madera" aumenta progresivamente al avanzar la sucesión.
c) Sustitución de unas especies por otras. Las especies pioneras u oportunistas colonizan de forma temporal los territorios no explotados. Se pasa de forma gradual de las especies “r estrategas”, adaptadas a cualquier ambiente, a especies “k estrategas”, mucho más exigentes y especialistas.r à k
d) Aumento en el número de nichos. Este incremento es debido a que, cuando se establecen relaciones de competencia, las especies “r” son expulsadas por las “k”, que ocupan sus nichos. El resultado final es una especie para cada nicho y un aumento en el número total de ellos.
e) Aumento de la diversidad. La comunidad clímax presenta una elevada diversidad que implica la existencia de un aumento del número de especies e interacciones.
f) Aumento de la respiración y disminución de la producción neta hasta equilibrarse en el clímax.
g) Evolución de los parámetros tróficos. La productividad decrece con la madurez. La comunidad clímax es el estado de máxima biomasa y mínima tasa de renovación (éste parámetro, también llamado productividad neta, nos da una idea de la velocidad de renovación de la biomasa de un ecosistema o nivel trófico. Puede variar entre 0-1, el uno sería el 100 % y se representa: r = PN/B)
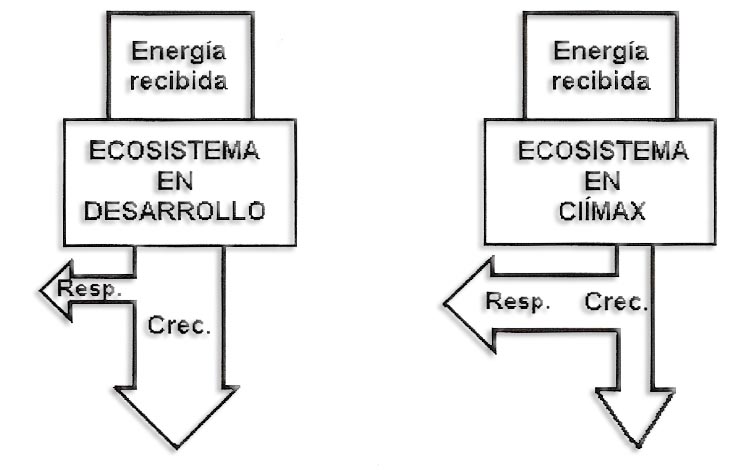
Si pensamos en la comunidad clímax de un ecosistema, la selva tropical es su máximo exponente: es un ecosistema cerrado pues la materia se recicla con gran rapidez por la eficaz acción de los descomponedores y se almacena en forma de biomasa.
5.2.1.2 CAUSAS DE LAS SUCESIONES
Pueden ser autógenas o exógenas.
a) Autógenas. Básicamente son modificaciones del microclima y del suelo. Si partimos de una etapa de pasto, con el tiempo se van implantando una serie de plantas leñosas cuya etapa final será un bosque.
Pasto à Matorral à Coscojar à Encinar
Esta sucesión sería típica de los suelos calizos mediterráneos.
La disminución de la luz que llega al suelo va produciendo la sustitución de especies heliófilas de las primeras etapas por otras umbrófilas.
b) Exógenas. Las más frecuentes son:
Grandes cambios climáticos. Como sucedió antes y después de las glaciaciones cuaternarias.
Cambios del microclima. Cuando se hace una tala o una entresaca masiva, se produce un aumento de la luz incidente en los niveles inferiores, lo que implica cambios de Tª y humedad que favorecen la presencia de plantas heliófilas.
Transformación del suelo. Si se elimina la vegetación clímax, la erosión aumenta y se asienta una vegetación xerofítica y pobre. Es el caso de las zonas mediterráneas donde se ha destruido el bosque esclerófilo mediterráneo (encinar) y que presentan un avanzado estado de desertización.
Influencia de la fauna. Animales como las cabras, pueden considerarse como agentes erosivos de primera magnitud. Igual ocurre cuando hay un exceso de herbívoros como conejos, gamos o ciervos en algunos cotos de caza.
Acción humana. A través de los siglos, el hombre ha alterado las sucesiones mediante talas, incendios, usos agrícolas, ocupación urbana e industrial, contaminación...
5.2.2 CLÍMAX
De forma general se puede decir que toda sucesión tiende hacia un estado final en equilibrio. Este estadio final hipotético recibe el nombre de clímax.
En condiciones normales, la situación clímax vegetal de nuestras latitudes es el bosque. Esta comunidad terminal estaría caracterizada por un gran número de especies vegetales y animales que son de mayor tamaño y tienen ciclos vitales más largos y complejos que los de las comunidades transitorias. También se aprecia una mayor cantidad de nichos ecológicos y de relaciones entre las especies.
Otras características son un máximo reparto espacial (estratificación) de las condiciones del medio, un mejor aprovechamiento de los nutrientes, un aumento drástico de la biomasa total de la comunidad y una relación producción bruta / respiración igual o próxima a la unidad.
De éste modo el clímax tiene una estabilidad e independencia máximas frente a los cambios ambientales.
La comunidad en el estado clímax continúa hasta que algún cambio origina una destrucción total o parcial de la misma, que ocasiona un desplazamiento de esta. Una destrucción total puede ser originada por una erupción volcánica, por una erosión intensa..., mientras que un incendio, generalmente suele determinar una destrucción parcial de la comunidad. La recuperación de la comunidad clímax depende de la magnitud de la transformación que haya sufrido. Si ha sido muy intensa, puede no haber recuperación.