



4. Respuesta humoral.
4. Respuesta humoral.
Es la respuesta inmunológica especifica producida por factores que se encuentran en el plasma y liquido intersticial, y que como hemos dicho antes los responsables son los linfocitos B.
4.1. Concepto, estructura y tipos de anticuerpos.
Los anticuerpos son proteínas globulares con una pequeña parte glucídica, que producen los linfocitos B, como respuesta especifica contra los antígenos.
Por sus propiedades inmunológicas reciben también el nombre de inmunoglobulinas (Ig).
Los anticuerpos o inmunoglobulinas (Ig) están formadas por 4 cadenas polipeptídicas, dosmás largas, llamadas por ello cadenas pesadas o cadenas H, y dos más cortas (cadenas ligeras o cadenas L), apareadas de tal modo que la molécula consta de dos mitades cada una de las cuales está constituida por una cadena larga y otra corta, adoptando el conjunto la forma de Y. La unión entre las cadenas se establece por puentes disulfuro (-S-S-) .Esta disposición permite distinguir en las Ig tres fragmentos moleculares: la denominada subunidad Fc (pie de la Y) y las denominadas subunidades Fab (brazos de la Y). Los aminoácidos que forman los extremos de cada fragmento Fab, tanto de las cadenas pesadas (H) como de las ligeras (L), son muy variables (VH y VL) mientras que en el resto son constantes sea cual fuere la inmunoglobulina a la que pertenecen (CH y CL). Las partes variables tanto de las cadenas L como de las H son las que permiten el acoplamiento al antígeno y definen la especificidad de la inmunoglobulina, formando el llamado centro activo de la misma o paratopo, que se une con el determinante antigénico o epítopo.
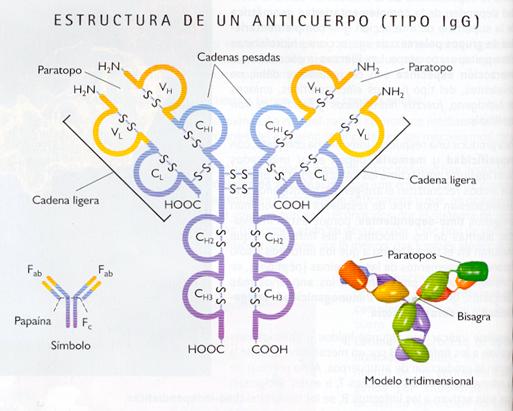
Los cinco tipos de inmunoglobulinas humanas se diferencian en la secuencia de aminoácidos de las cadenas H:
Ig G: es la más abundante. Aparece después de la Ig M en la respuesta humoral y permanece aún cuando ha desaparecido el antígeno. Atraviesa la placenta y proporciona defensas al recién nacido en las primeras semanas, neutraliza las toxinas, fija el complemento y opsoniza los microorganismos patógenos.
Ig M: es la primera en aparecer en la respuesta humoral y sólo se encuentra en sangre, de donde no puede salir debido a su elevado peso molecular. Son agentes aglutinantes y citolíticos, también actúan como receptores de membrana en los linfocitos B.
Ig A: aparece en el suero y en las secreciones seromucosas (saliva, lágrimas, fluidos nasales, calostros, etc.) donde actúan como defensas de las superficies externas del cuerpo. No se fijan al complemento.
Ig E: se localizan como receptores en las membranas de los mastocitos y de los basófilos; ante la presencia de un antígeno, que suele ser un alergeno, se produce la desgranulación del citoplasma y la liberación de histamina y otras aminas vaso activas responsables del asma y las alergias.
Ig D: permanece unida a las membranas de los linfocitos B, donde probablemente desempeña la función de receptor antigénico, junto con la Ig M.
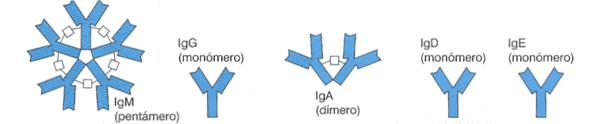
Estructura de los cinco tipos de inmunoglobulinas humanas
4.2. Células productoras de anticuerpos: linfocitos B.
Los linfocitos B se forman y diferencian en la médula ósea. Allí adquieren capacidad para producir anticuerpos, moléculas específicas contra los antígenos invasores. Por esta razón se dice que son inmunocompetentes.
En la médula ósea se generan millones de linfocitos B, genéticamente diferentes, cada uno de los cuales fabricará distintos anticuerpos, capaces de unirse a la enorme cantidad de antígenos diferentes que entran en un organismo.
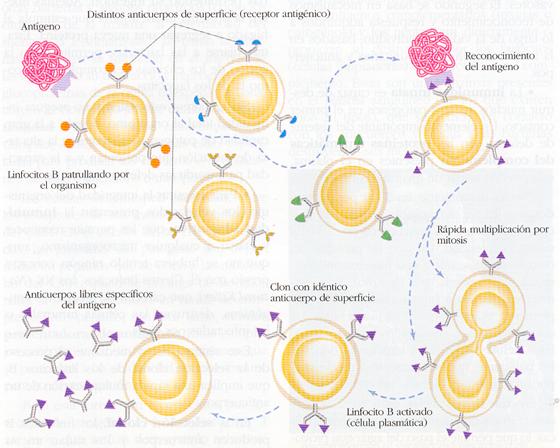
Por lo general, en el sistema inmunitario sólo hay unos pocos linfocitos B diferenciados que, además, se encuentran inactivos. Cuando aparece un antígeno, tiene lugar su unión a un anticuerpo de la membrana de un determinado linfocito B. Esta unión únicamente es posible si existe un acoplamiento espacial entre ambas moléculas; es decir, es necesaria una conformación complementaria entre la estructura del antígeno y la del anticuerpo. El reconocimiento antigénico se basa, por tanto, en la especificidad de la unión con el anticuerpo correspondiente. Dada la enorme variedad de anticuerpos, siempre existe alguno capaz de reconocer a un antígeno concreto. La unión puede ser fuerte o débil. En el primer caso, la respuesta inmunitaria generada es más intensa.
Tras el reconocimiento del antígeno, los linfocitos B se activan. Su activación consiste en la rápida división de estas células para originar una serie o clon de células iguales productoras del mismo tipo de anticuerpo. Así, además de aumentar considerablemente el número de linfocitos después de la exposición al antígeno, todos ellos generan los anticuerpos específicos correspondientes. De esta forma, la propia aparición del antígeno sería la causa de una selección clonal por la que se obtiene un número elevado de linfocitos B productores de los anticuerpos necesarios contra ese antígeno.
Esta teoría de la selección clonal , elaborada por F. M. Burnet, permite explicar por qué se producen grandes cantidades de anticuerpos específicos tras la introducción de un determinado antígeno. Es el mismo antígeno el que, por su unión específica a un linfocito concreto, de entre los millones existentes, induce a éste a originar, a partir de él, un clon encargado de rechazarlo.
Los linfocitos que no contactan con el antígeno específico no se activan, pero siguen disponibles por si éste aparece en el futuro.
La mayor parte de los linfocitos B activados se convierten en las llamadas célulasplasmáticas , de gran tamaño y con una enorme producción de anticuerpos (unas 10000 moléculas/célula/minuto). Sin embargo, algunos de ellos quedan como linfocitos B de memoria, que tienen una vida ilimitada y constituyen una reserva inmunitaria para nuevas exposiciones futuras al mismo antígeno.
Tanto las células plasmáticas como las de memoria se acumulan en la zona cortical de los ganglios linfáticos, desde donde pasan a la linfa y de ésta a la sangre.
La activación de los linfocitos B se intensifica mucho por la acción de las interleucinas, unas sustancias segregadas por ciertos linfocitos T y por los macrófagos.

4.3. Reacción antígeno-anticuerpo.
Tanto la respuesta humoral como la celular suponen el reconocimiento de determinadas estructuras químicas en la superficie de macromoléculas extrañas, los antígenos. Estas estructuras químicas del antígeno se denominan determinantes antigénicosy la especificidad de la respuesta inmunitaria va dirigida especialmente a ellos.
Cuando se ponen en contacto un antígeno con el anticuerpo específico, reaccionan uniéndose mediante un enlace no covalente entre la zona específica de la inmunoglobulina y los determinantes antigénicos de la molécula de antígeno. La combinación del anticuerpo con el antígeno desencadena una serie de procesos capaces de neutralizar y eliminar a una sustancia extraña.
Las reacciones más importantes entre antígeno y anticuerpo son las siguientes:
I. Precipitación. Se produce entre antígenos y anticuerpos solubles que al unirse forman agregados insolubles de ambas moléculas que precipitan.
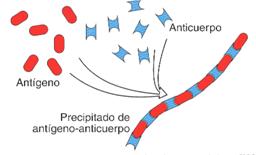
Mecanismo de precipitación de un antígeno soluble y un anticuerpo
2. Aglutinación. En este caso, los anticuerpos se dirigen contra los antígenosque se encuentran en lasuperficie de ciertas célulascomo microorganismos o glóbulos rojos. El anticuerpo se combina con los antígenos de superficie y forma aglomerados de células.
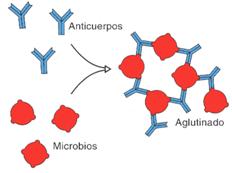
Aglutinación
La hemaglutinación se produce entre los anticuerpos del plasma sanguíneo y los antígenos de los glóbulos rojos de la sangre de diferente grupo sanguíneo. Esta prueba es fundamental en la determinación de los grupos sanguíneos ABO.
3. Neutralización. La unión de los receptores específicos del anticuerpo al antígeno bloquean la acción de éstos contra las células de los tejidos invadidos.
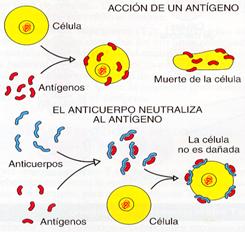
Neutralización de un antígeno por un anticuerpo específico
4. Opsonización. La llevan a cabo anticuerpos que reciben el nombre de opsoninas. Las opsoninas se unen a antígenos presentes en superficies celulares de bacterias y forman un revestimiento que favorece la fagocitosis por los macrófagos. Estos anticuerpos hacen posible la acción fagocitaria que sin ellos sería imposible.
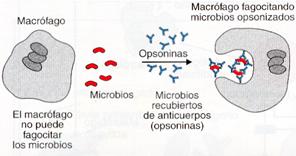
Las opsoninas favorecen la fagocitosis de los microbios