



1.2. Grupos principales.
1.2. Grupos principales.
Como hemos dicho anteriormente los microorganismos pertenecen a distintos grupos taxonómicos y para clasificarlos vamos a seguir la clasificación de Whittaker (1969) modificada por Margulis, que establecía la existencia de cinco reinos, de los cuales a sólo tres pertenecen los microorganismos, además, en los microorganismos se incluyen también formas que no pertenecen a ningún reino por ser acelulares, de este modo la clasificación la podemos hacer de la siguiente manera, teniendo en cuenta su organización y otras características y así tenemos:
- Formas acelulares: - No tienen metabolismo: Virus y similares
(viriones, priones…)
- Formas celulares: - Organización procariota
§ Reino Monera
Ø Arqueobacterias
Ø Eubacterias o bacterias
- Organización eucariota
§ Reino Protista
Ø Algas
Ø Protozoos
§ Reino Fungii
Ø Hongos
Estos grupos se dividen en otros, que podemos ver en los siguientes cuadros:
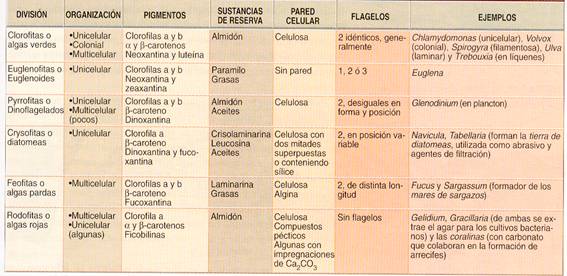
Clasificación y características de las algas
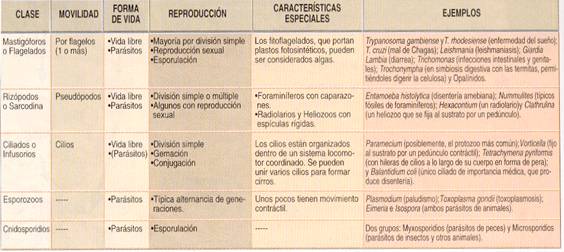
Clasificación y características de los protozoos
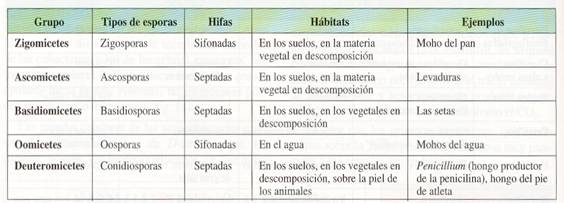
Clasificación y características de los hongos
1.2.1. Formas acelulares: virus.
Los virus son los organismos más simples y pequeños que se conocen, pertenecen a un nivel de organización acelular, y marcan la barrera entre lo vivo y lo inanimado. Son parásitos intracelulares obligados que alteran en su propio beneficio los mecanismos biosintéticos de las células hospedadoras con el fin de lograr su reproducción. Son fragmentos de ácido nucleico cubiertos por una envoltura proteica que les permite desplazarse de una célula a otra, comportándose como elementos genéticos móviles. Se les puede considerar de dos maneras, como agentes transmisores de enfermedades y como agentes trasmisores de herencia. En las enfermedades víricas alteran las funciones celulares pudiendo llegar a ocasionar la muerte de la célula hospedadora. Como transmisores de herencia el ácido nucleico viral puede permanecer en la célula hospedadora y producir cambios genéticos perjudiciales o beneficiosos.
1.2.1.1. Composición, estructura y actividad biológica.
Los virus pueden alternar entre dos estados distintos, intracelular y extracelular.
En el estado extracelularla partícula viralo viriónes inerte metabólicamente, siendo su única función transportar el ácido nucleico viral desde la célula en la que se ha reproducido hasta otra célula en la que se pueda reproducir. Según S. Luria, actúan como "pedacitos de herencia en busca de cromosomas". La morfología de los virus se refiere a este estado, en el que constan de ácido nucleico, cubierta y enzimas.
Un solo tipo de ácido nucleico, ADN o ARN, existiendo virus con modalidades de ácidos nucleicos no frecuentes, como ADN monocatenario y ARN bicatenario. Este genoma codifica básicamente enzimas para la replicación de su ácido nucleico. Puede estar formando una sola molécula o fragmentado en varias. Así, el virus del SIDA tiene dos moléculas idénticas de ARN, y el de la influenza consta de ocho fragmentos de ARN. Los virus difieren en el tamaño, cantidad y características de su ácido nucleico.
Una cubierta proteicao cápsidaque aloja en su interior al ácido nucleico. Tiene como misión proteger al ácido nucleico y reconocer los lugares de unión y penetración de las células a parasitar, en aquellos virus que carecen de membranas externas. Está formada por repetición de proteínas globulares llamadas capsómeros. La variedad de estos capsómeros es baja, aunque su número sea elevado. Estos capsómeros se autoensamblan según una información contenida en ellos mismos, originando figuras simétricas, por lo que los virus pueden ser estructuras cristalizables. Al conjunto de cápsida y ácido nucleico se le conoce como nucleocápsida. Según su simetría, los virus pueden ser:
a) Virus de simetría helicoidal. Los capsómeros se disponen helicoidalmente, sobre la hélice de ácido nucleico, adquiriendo globalmente la forma de bastoncillo. A este tipo pertenece el virus del mosaico del tabaco y el virus de la rabia.
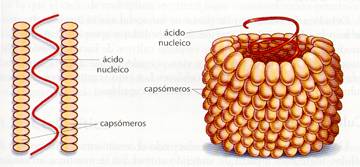
Esquema de un virus helicoidal
b) Virus de simetría icosaédrica. Adoptan la forma de un poliedro de 20 caras triangulares, obtenidas por la asociación de varios tipos de capsómeros. En conjunto tienen un aspecto casi esférico. Pueden tener proteínas fibrilares que salen del icosaedro. Ejemplos de esta modalidad son los adenovirus y el virus de la polio.
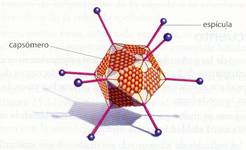
Esquema de un virus con simetría icosaédrica
c) Virus complejos. Compuestos de varias partes diferentes, de forma y simetría distinta. Los bacteriófagos son ejemplos de este grupo. Constan de cabeza icosaédrica donde se aloja el ácido nucleico, cola helicoidal que se puede contraer para inyectar el contenido de la cabeza, estando separados por un collar o cuello formado por capsómeros distintos. Al final de la cola está la placa basal, especializada en la fijación sobre la célula hospedadora. De ella parten unas fibras que potencian esta fijación. Estos virus deben tener más cantidad de ácido nucleico que otros virus, pues necesitan codificar varios tipos de capsómeros.
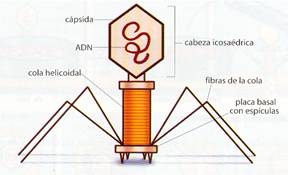
Esquema de un virus complejo: bacteriófago T4
d) Virus envueltos. Tienen rodeada la nucleocápsida por estructuras membranosaslipoproteicas, que son fragmentos de la membrana de la célula hospedadora en la que se llevó a cabo la reproducción viral. De estas membranas pueden partir espículas, con capacidad enzimática, normalmente codificadas por el propio virus. Los virus que no tienen estas membranas, y sólo tienen nucleocápsida se conocen como virus desnudos.
|
A |
|
B |
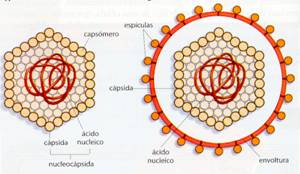
Esquema de un virus desnudo (A) y con envoltura (B)
Enzimas virales.Aunque los viriones no tienen metabolismo fuera de la célula hospedadora, es decir, son inertes, pueden contener enzimas que actúan en el proceso infeccioso, ya sea en la entrada o salida de las células en las que se han reproducido o en la replicación o transcripción de su genoma.
Como vemos, los virus carecen de citoplasma, de orgánulos y de enzimas con los que realizar metabolismo alguno, y la única función vital que realizan es la reproducción dentro de una célula viva.
En el estadointracelular el ácido nucleico se integra en la célula hospedadora. El virus se reproduce, replicando su genoma y sintetizando los componentes de la cubierta del virus. Su "aspecto" está en función de la fase del ciclo reproductor en la que se encuentre.
1.2.1.2. Ciclos de vida de los virus: lítico y lisogénico.
En el estado extracelular los virus son inertes, no presentan metabolismo y no requieren materia ni energía, sólo " esperan" encontrar a la célula hospedadora sobre la que se puedan fijar e introducir su genoma para llevar a cabo la reproducción viral.
En el estado intracelular el genoma vírico es capaz de inhibir la expresión génica de la célula hospedadora y dirigir la maquinaria metabólica de dicha célula hacia la producción de copias del ácido nucleico viral (replicación) y de las proteínas de su cápsida o enzimáticas (transcripción y traducción), con el fin de producir nuevas partículas vírales, que tras salir de esa célula puedan infectar a otras células.
Dentro de los ciclos de reproducción víricos podemos distinguir cinco fases, a) fijación; b) penetración; c) eclipse; d) ensamblaje y e) liberación, pudiendo retrasarse las tres últimas fases en los ciclos lisogénicos que explicaremos posteriormente.
a) Fase de fijación o adsorción. Las células hospedadoras tienen en sus membranas receptores específicos para los virus que las infectan, uniéndose componentes de la cápsula o membranas externas del virus con glucoproteínas o lipoproteínas concretas de la membrana de la célula infectada. La unión puede ser mecánica, como la realizada por las espinas basales de los bacteriófagos, o con más frecuencia por enlaces químicos. Estos receptores realizan funciones normales en las células y no se encuentran en todas las células de un organismo. A lo largo de la evolución cada tipo de virus ha adquirido la capacidad de fijarse a receptores específicos en la membrana de un determinado tipo celular. Por ejemplo algunos bacteriófagos se fijan a los pelos bacterianos, el virus de la gripe se une a glucoproteínas de las membranas de las células de las mucosas, y el receptor de la polio es una lipoproteína de las membranas del tejido nervioso. Si estos receptores faltan o se modifican, por mutación o por fármacos, el hospedador puede hacerse resistente a la infección al no fijarse el virus, pero también pueden aparecer mutantes del virus capaces de adherirse a hospedadores resistentes.
En muchos virus vegetales no se han localizado estos receptores.
b) Fase de penetración. En función de las estructuras superficiales de la célula hospedadora los virus utilizan distintos mecanismos de penetración.
- Los bacteriófagos, como el T4 de E.coli, originan un pequeño orificio en la pared bacteriana con la lisozima de su placa basal, por el que se inyectael ADN, al contraerse la cola del virus. Este ADN debe estar muy empaquetado pues tiene una longitud 500 veces superior al de la cabeza que lo contiene. La cápsida queda en el exterior de la bacteria.
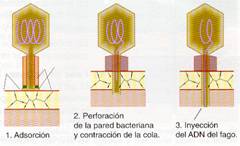
Inyección del ADN vírico del bacteriófago T4 de E.coli
- Los virus desnudos introducen toda la nucleocápsida en la célula, ya sea por penetración directa al perforar la membrana con enzimas hidrolíticos, o por endocitosis, al ser englobados por la célula hospedadora en una vacuola, que romperán dentro de la célula.
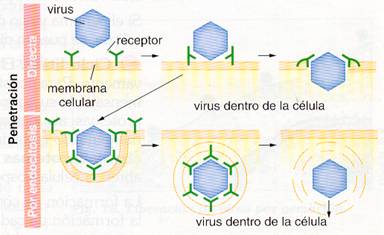
Mecanismos de penetración de virus desnudos
- Los virus envueltos funden su cubierta con la membrana de la célula hospedadora, proceso comprensible pues son de la misma naturaleza, como veremos en la fase de liberación. Puede ser una fusión directa o aplazarse, entrando el virus por endocitosis dentro de una vacuola, de la que es liberado el virus al producirse la fusión con un lisosoma celular. En ambos casos burlan la protección que supone para la célula hospedadora su membrana celular.
En estas dos últimas modalidades en las que penetra la nucleocápsida, ésta sufre una separación de sus capsómeros o desensamblaje (descapsización), con la liberación del ácido nucleico y de las escasas enzimas que pudiera haber en la cápsida.
En todos los casos se necesitan muchas partículas víricas para que una consiga penetrar en la célula hospedadora.
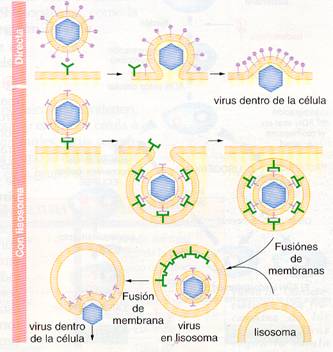
Penetración por fusión directa o con lisosomas
c) Fase de eclipse. En ella no se detectan virus en el interior de la célula. El genoma vírico dirige el metabolismo de la célula hospedadora hacia la síntesis de los componentes víricos, utilizando todos los recursos de la célula hospedadora (materias primas, nucleótidos y aminoácidos, energía o ATP, ARN transportadores, ribosomas y enzimas).
Como este genoma vírico es extraño a la célula puede ser atacado por los enzimas de restricción y modificación de la célula hospedadora, que reconocen al ADN extraño, siendo éste otro medio de defensa contra la infección vírica. Penetración no siempre implica infección.
Si el genoma vírico no es destruido, en función de la duración de la fase de eclipse, se pueden distinguir dos ciclos víricos: Ciclo lítico y ciclo lisogénico.
Ciclo lítico. El virus se apropia de la célula para multiplicarse masivamente. Para construir los nuevos virus, es necesario que se transcriba el mensaje de su genoma en ARN mensajeros, y la traducción de éstos en proteínas; algunas enzimáticas, que controlaran la replicación de su material genético (proteínas o enzimas tempranas), otras que formarán la cápsida (proteínas tardías) y las últimas son las enzimas líticas, que abren la célula hospedadora y liberan a los nuevos virus.
La formación de copias de su genoma, replicación, se realiza mediante la formación de cadenas complementarias. En los casos de ARN o ADN bicatenarios, cada cadena actúa como molde de la complementaria. Los virus ADN monocatenarios sintetizan inicialmente otra cadena complementaria, replicándose a partir de este momento como los bicatenarios.
La transcripción y traducción en proteínas se realiza tras la replicación utilizando todos los componentes de la célula hospedadora necesarios para esta función.
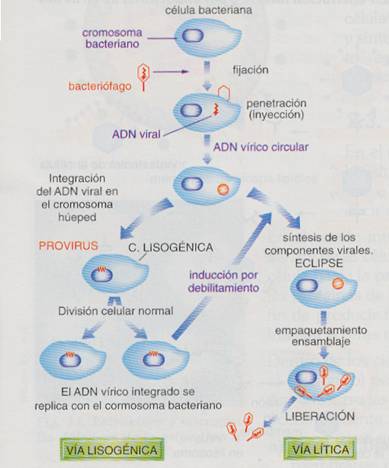
Sucesión de ciclos líticos o lisogénicos
Ciclo lisogénico. Tras la penetración del cromosoma viral no siempre es inmediata la multiplicación del virus. Muchos virus entran en un estado de latencia, posponen su reproducción. El cromosoma viral puede integrarse en algún cromosoma de la célula hospedadora, caso más frecuente, o quedar en el citoplasma, a modo de plásmido. A este estado del virus se les conoce como virus atenuadoso provirus, ya la célula que los aloja como célula lisogénica. La célula lisogénica, que transporta el provirus, puede multiplicarse indefinidamente durante muchas generaciones, hasta que por agresiones ambientales que causan el debilitamiento de la célula, se induce al provirus a dejar el cromosoma hospedador y comenzar un ciclo normal o lítico de reproducción. De esta manera el provirus no perece con la célula, sino que forma nuevos virus que infectaran a otras células. Este proceso significa un enriquecimiento y alteración genética de la célula hospedadora, con graves consecuencias como infecciones latentes, mutaciones, transformaciones cancerosas. Pero también por recombinación génica puede estar ligado a procesos de evolución de las especies. Ejemplos de virus lisogénicos son muchos bacteriófagos, los retrovirus, el virus de las verrugas y el del herpes.
d) Fase de ensamblaje. Se ensamblan los capsómeros formando la cápsida, a la vez que el ácido nucleico se pliega en su interior, junto a las enzimas que pueda llevar el virus. Este ensamblaje suele ser automático, basado en la estructura de los propios capsómeros, pero también pueden intervenir enzimas de origen viral.
e) Fase de liberación. Los virus salen de la célula básicamente por dos mecanismos:
-Por gemación, induciendo en la membrana celular la formación de pequeñas vesículas en las que se introducen, y que acaban separándose de la célula. Este procedimiento lo utilizan los virus envueltos, en los que la envoltura membranosa es parte de la membrana de la célula en la que se reprodujeron.
-Aprovechando los mecanismos de exocitosis de la célula, o provocando pequeños agujeros en su membrana por medio de sus enzimas líticos. Los virus desnudos usan estos mecanismos.
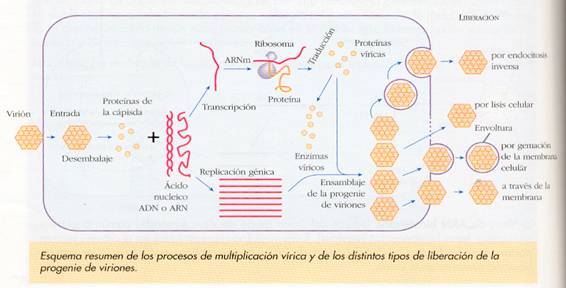
Esta liberación de los virus puede causar la muerte de la célula hospedadora, ya sea por haber agotado sus nutrientes, roto su membrana si la liberación ha sido masiva, o destruido su genoma. Este es el ciclo líticopropiamente dicho. En otros casos, la liberación puede ser lenta, sin lisar a la célula, la cual puede seguir viva y producir virus durante un tiempo variable. Este caso se conoce como infección persistente. Cuando los virus son lisogénicos, como la reproducción está retardada, se conoce como infección latente.
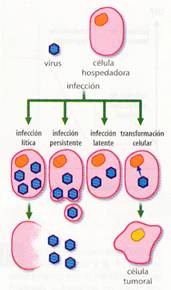
Distintos tipos de infecciones víricas
Un grupo muy especial de virus animales son los RETROVIRUS.Retro significa hacia atrás, recibiendo este nombre porque, siendo virus ARN, se replican por medio de un intermediario de ADN, en sentido contrario al ADN. Son virus envueltos, con simetría incierta y varios tipos de proteínas en su cubierta. Su genoma consta de dos moléculas iguales de ARN monocatenario, y al menos tienen 3 tipos de enzimas: transcriptasa inversa, ADN endonucleasa o integrasa, y una proteasa que favorece la salida del virus.
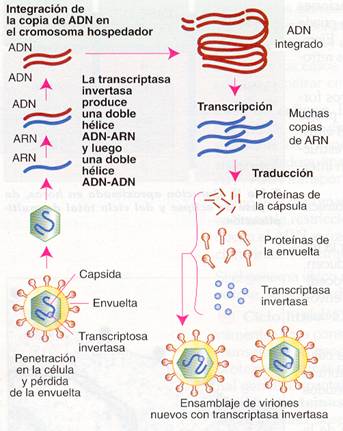
Replicación de un retrovirus
1.2.2. Formas celulares: bacterias (procariotas) y algas, protozoos y hongos (eucariotas).
Aquí se incluyen los microorganismos que tienen una organización celular, que puede ser procariota, es decir, sin un verdadero núcleo, como las bacterias, o una organización eucariota con verdadero núcleo que tienen las algas, los protozoos y los hongos.
1.2.2.1. Características estructurales.
Vamos a estudiar las características estructurales de cada grupo por separado, porque a pesar de tener la organización típica de las células procariotas y eucariotas, tienen grandes diferencias.
BACTERIAS:
Las bacterias son microorganismos unicelulares, de dimensiones comprendidas entre 1 micra y 5 micras de diámetro. Pueden habitar en todos los medios terrestres y acuáticos, e incluso en el interior de otros seres vivos. La forma de las bacterias es muy diversa; las más características son los bacilos que tienen forma de bastón, los cocos que tienen forma esférica, los espirilos que parecen un tirabuzón y los vibrios, que tienen forma de coma.
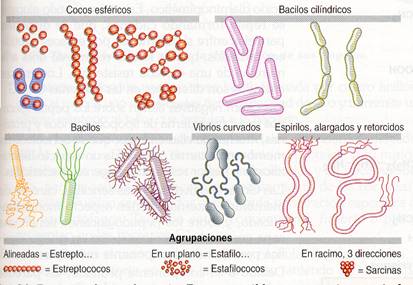
Formas posibles y agrupaciones más frecuentes de bacterias
Estructura de la célula bacteriana:
La estructura de una célula bacteriana corresponde a la de procariota.
El estudio detallado al microscopio óptico y al microscopio electrónico ha permitido distinguir su estructura.
Las células bacterianas están provistas de pared celular, membrana plasmática, citoplasma, cromosoma o genóforo e inclusiones citoplasmáticas, como los ribosomas. Además, algunas poseen estructuras superficiales como cápsulas, flagelos y fimbrias.
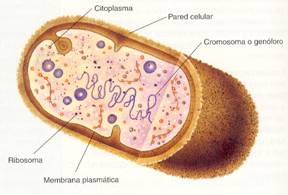
Estructura de una bacteria
· La pared celular es una cubierta rígida que recubre la membrana plasmática. Está constituida principalmente por mureína, un peptidoglicano formado por la unión de monómeros de N-acetilglucosamina, y ácido N-acetilmurámico, moléculas que se unen por medio de enlaces O-glucosídicos. Estas moléculas se disponen formando una malla que envuelve la célula.
Existen dos tipos de pared que difieren en su grosor y forma. Fueron descubiertas por Hans Christian Gram en 1884, el cual comprobó que podían diferenciarse según un método de tinción, que fue denominado tinción de Gram en su honor. Así, podemos diferenciar:
- Bacterias Gram positivas, que se tiñen de color violeta. Presentan una pared de unos 50 nm de grosor, de estructura lisa.
- Bacterias Gram negativas, que se tiñen de color rojo. La pared es más delgada, de unos 10 nm de grosor, y su estructura externa es irregular.
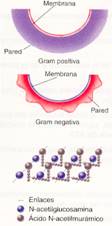
Tipos de bacterias según el método de tinción
· La membrana plasmática tiene una estructura similar a la de la célula eucariota, con un espesor de 7,5 a 8 nm. Sin embargo, difiere en algunas de sus funciones.
Está formada por una doble capa de fosfolípidos, con proteínas incluidas. Los fosfolípidos presentan las regiones hidrófilas polares hacia el exterior de la membrana y hacia el citoplasma. Las cadenas hidrófobas apolares de ácidos grasos se dirigen hacia el centro de la bicapa.
La membrana contiene múltiples proteínas, entre ellas enzimas como permeasas, y, en algunos grupos, pigmentos fotosintéticos, como bacterioclorofilas.
En muchas especies, puede formar pliegues hacia el interior de la célula. De este modo, se aumenta su superficie. Estos pliegues se denominan mesosomas.
· Las bacterias contienen un único cromosoma, denominado genóforo. No posee envoltura nuclear que lo separe del resto de la célula. El cromosoma está constituido por una doble cadena de ADN, cerrada sobre sí misma y muy replegada.
En ocasiones, también contienen plásmidos, que son un fragmento circular de ADN bicatenario extra que existe en algunas bacterias como material con capacidad replicativa autónoma.
· El citoplasma tiene una estructura fibrosa, similar al citoesqueleto de las células eucariotas. Está constituido por proteínas que se entrelazan para formar una matriz. El resto está formado por un 80 % de agua.
Además del genóforo, contiene los ribosomas y las inclusiones citoplasmáticas. Las fibrillas entrelazan todos los componentes del interior de la célula, incluidos los ribosomas, que en la célula eucariota están libres.
En todas las formas bacterianas se encuentran ribosomas y otras inclusiones características.
· Los ribosomas tienen un tamaño algo más pequeño que los ribosomas de las células eucariotas, son 70S, con subunidades 50S y 30S.
· En las células bacterianas existen otras inclusiones rodeadas por una membrana distinta a la plasmática. Este tipo de membrana consiste en una única capa en la que predominan las proteínas, y cuyo tamaño oscila entre 2 y 5 nm de grosor.
- Las vacuolas de gas son vesículas de aspecto cilíndrico. Favorecen la flotación de las bacterias que viven en un medio acuático.
- Los carboxisomas, que también se denominan cuerpos poliédricos, son pequeñas estructuras membranosas que contienen ribulosa difosfato carboxilasa (rubisco), el principal enzima fijador de dióxido de carbono para el ciclo de Calvin.
Por tanto, están presentes en las bacterias fotoautótrofas y quimioautótrofas.
- Los clorosomas son vesículas alargadas que tapizan el interior de la membrana plasmática y contienen pigmentos verdes fotosintéticos.
Son propios de las bacterias que realizan la fotosíntesis.
- Otras inclusiones contienen materiales celulares de reserva.
Así, se presentan diversos tipos de inclusiones citoplasmáticas, como las de almidón, glucógeno, ácido 1, 3-hidroxibutírico o polifosfatos.
Además existen otras estructuras que pueden pertenecer a determinados tipos de bacterias.
Entre estas estructuras encontramos: cápsula, flagelos y fimbrias
· La cápsula es una envoltura de grosor variable y de aspecto mucoso que puede formarse en algunas bacterias por la parte externa de la pared celular. Está formada por cadenas de polisacáridos.
· Los flagelos son filamentos semirrígidos, presentes en algunas especies de bacterias, cuya función consiste en proporcionar movimiento a las células.
El número y la posición de los flagelos varían en las distintas especies, y se pueden clasificar las bacteria flageladas como monotricas (con un flagelo en un extremo), lofotricas (dos o más en un extremo), anfitricas (flagelos en ambos extremos) y peritricas (flagelos en toda la superficie bacteriana)
|
|
|
|
|
Tipos de bacterias según su número y posición de los flagelos |
||
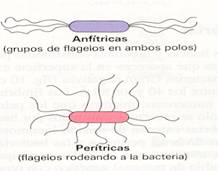
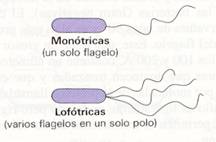
Los flagelos parten de la membrana plasmática, atraviesan la pared y se extienden hacia el exterior de la célula.
La forma y el diámetro también pueden cambiar. Este último oscila entre 12 y 18 nm.
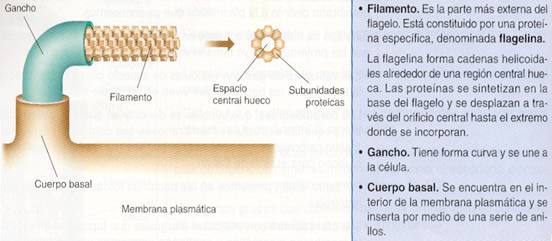
Estructura de un flagelo
· Las fimbrias o pili son filamentos rígidos, formados por agrupaciones en forma helicoidal de la proteína pilina. Su grosor es muy variable, pueden medir desde 4 nm de diámetro hasta 35 nm.
Las fimbrias se clasifican según su función:
- Fimbrias de infección. Las poseen bacterias que parasitan otras células. Permiten la adhesión a las células que infectan.
- Fimbrias sexuales. Facilitan el proceso de la conjugación, es decir, la transferencia de material genético de una célula a otra.
ALGAS:
Tienen organización eucariota, con cloroplastos que contienen distintos pigmentos fotosintéticos además de las clorofilas y carotenoides, tales como ficobilinas (ficocianina y ficoeritrina) o fucoxantinas. Son algunos de estos pigmentos los que confieren el color característico algunos grupos.
La mayoría tienen pared celular, cuyo polímero fundamental es la celulosa, otros grupos carecen de pared (Euglenofitas) y otras como las diatomeas tienen un caparazón de sílice organizado en dos valvas.
Los polímeros carbonados de reserva son muy variados y sólo algunos sintetizan almidón.
Pueden tener flagelos o no.
Las algas son organismos o bien unicelulares, bien coloniales, o bien pluricelulares.
Para ver mejor su estructura vamos a ver dos ejemplos, como son la Euglena y Volvox, esta última es una colonia formada por cientos de Chlamydomonas.
|
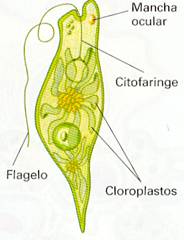
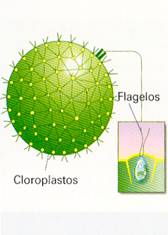
EUGLENA La célula es alargada. Presenta numerosos cloroplastos. En uno de sus extremos la membrana plasmática se pliega hacia dentro y forma una especie de saco, llamado citofaringe, del cual salen dos flagelos. Uno de estos flagelos es muy corto. Cerca de la citofaringe se encuentra una mancha roja de forma ovalada llamada mancha ocular. Esta mancha es sensible a la luz y percibe su intensidad y dirección. Gracias a ello, la Euglenase desplaza hacia las zonas donde puede captar suficiente luz para realizar la fotosíntesis. Estas algas viven en las aguas dulces y en el lodo de lagos, estanques o ríos. Abundan principalmente en aguas estancadas, a las que dotan de un característico color verde. VOLVOX El alga denominada Volvox está constituida por cientos o miles de células, todas muy parecidas entre sí. Esta agrupación de células se denomina colonia. En las colonias, las células se distribuyen formando una esfera hueca, de aproximadamente un milímetro de diámetro, recubierta por una capa gelatinosa que las mantiene unidas. Cada célula tiene las siguientes características: Presenta un gran cloroplasto para realizar la fotosíntesis. Posee dos flagelos situados hacia el exterior, con los que contribuye al movimiento de la colonia. Tiene una mancha ocular sensible a la luz. El Volvox es un alga muy frecuente en aguas dulces. |
|
PROTOZOOS:
Es un grupo muy heterogéneo, con estructura unicelular, aunque algunos grupos forman colonias con un número variable de individuos. Existen colonias con células sin ningún tipo de especialización, pero también hay especies coloniales en las que se diferencian células vegetativas y células reproductoras.
Por otro lado, numerosos ejemplares presentan características de los seres vegetales, como la presencia de pigmentos sensibles a la luz y la capacidad para realizar la fotosíntesis.
La estructura celular que presentan los protozoos son las características de la célula eucariota.
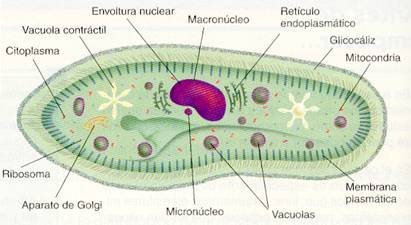
Estructura celular de un protozoo
Membrana plasmática. La célula está rodeada por una membrana plasmática típica, es decir, una bicapa lipídica con proteínas incluidas.
En algunos grupos de protozoos, por la parte exterior de la membrana, se puede presentar un glicocáliz, compuesto de mucopolisacáridos.
Citoplasma. Se pueden distinguir el hialoplasma y los orgánulos rodeados de membrana
En el hialoplasma es habitual diferenciar una parte fluida y homogénea y otra más viscosa, de aspecto granular y con abundantes inclusiones.
Respecto a los orgánulos, podemos encontrar los componentes característicos de cualquier célula eucariota animal con sus funciones habituales.
Son comunes a todos los grupos tanto el aparato de Golgi como el retículo endoplasmático, aunque pueden tener extensión variable según las características del individuo y el estado fisiológico de la célula.
Enalgunos grupos se dan modificaciones en la estructura básica como consecuencia de su modo de vida.
- Los protozoos parásitos presentan muy pocas mitocondrias y en algunos casos ninguna.
- Es muy frecuente que las células tengan unas formaciones rodeadas de membrana situadas en la periferia de la célula. Por acción de diversos estímulos, estas estructuras vierten su contenido al exterior.
Su función es todavía indeterminada, aunque se relacionan con procesos como la constitución de cubiertas esqueléticas exteriores a la célula o la captura y digestión de presas.
- La mayoría de los protozoos que viven libres en las aguas dulces presenta vacuola contráctil. Este orgánulo permite evacuar al exterior el agua que, por fenómenos de ósmosis, penetra en el interior de la célula.
Aparte de los orgánulos, en el citoplasma se encuentran inclusiones formadas por materiales de reserva, como polisacáridos y lípidos, y materiales de excreción diversos.
Núcleo. Varía en cuanto a forma y número según los distintos grupos. En general, los protozoos presentan células uninucleadas, aunque existen casos de células de gran tamaño que contienen muchos núcleos.
Algunas especies tienen dos núcleos: un macronúcleoo núcleo grande y un micronúcleo o núcleo pequeño. Aparte de su tamaño, estos núcleos presentan diferencias estructurales y funcionales:
- El micronúcleo es diploide y no produce ARN.
- El macronúcleo es poliploide y produce el ARN necesario para la síntesis de proteínas de la célula.
Cilios y flagelos. Se presentan en las especies que se desplazan activamente.
En numerosos casos, los protozoos poseen elementos esqueléticos que defienden al organismo de agentes exteriores perjudiciales y contribuyen a mantener su forma. Pueden ser internos o externos y en algunos casos, más que un esqueleto, se trata de una especie de concha donde el organismo vive con independencia de esta cubierta.
Por otro lado, muchas especies presentan formas de resistencia como quistes, recubiertas por una capa extraordinariamente resistente.
HONGOS:
Se suele denominar hongos a un grupo muy heterogéneo de seres vivos. Para centrar el tema de esta parte de la unidad, diferenciamos distintos tipos de organismos incluidos en este grupo.
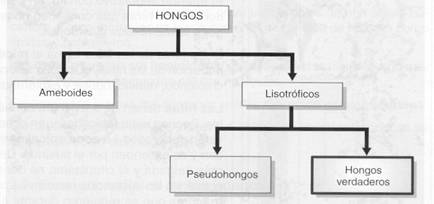
Todos los hongos están constituidos por células eucariotas y son heterótrofos. Las diferencias entre los distintos tipos se establecen, básicamente, por su estructura y el modo en que captan el alimento.
Los hongos ameboides son un grupo de hongos cuya estructura recuerda a la de las amebas. Están formados por células sin pared y plurinucleadas; la mayoría de las especies capta el alimento por fagocitosis.
El resto de los organismos conocidos como hongos corresponde al grupo de los hongos lisotróficos.
Éstos tienen una estructura filamentosa, con pared celular exterior a la membrana plasmática.
Son heterótrofos pero, a diferencia de los hongos ameboides, realizan digestiónexternao lisotrofía. Segregan las sustancias necesarias para la digestión del alimento y luego absorben los nutrientes resultado de la digestión.
Aunque aparentemente se observa una considerable semejanza entre todos los hongos lisotróficos, se pueden distinguir dos grupos: los pseudohongos y los hongos verdaderos.
Los pseudohongos se caracterizan por la presencia de celulosa como componente más abundante de la pared celular, mientras que en los hongos verdaderos el componente más abundante es la quitina.
El objetivo de esta parte de la unidad es el estudio detallado de los hongos verdaderos, un grupo de seres vivos que la mayoría de los especialistas considera un reino propio, el reino hongos.
En los hongos verdaderos se observa una evolución progresiva hacia la adaptación a la vida terrestre.
Iniciamos el estudio de los hongos verdaderos describiendo su estructura.
Entre los hongos verdaderos se puede distinguir una estructura miceliar y una estructura levuriforme.
Estructura miceliar
El aparato vegetativo consta, en general, de filamentos alargados denominados hifas que contienen numerosos núcleos y una pared exterior a la membrana plasmática.
El conjunto de las hifas forma el micelio. Gracias al gran poder de penetración de las hifas, el micelio se encuentra en permanente actividad, creciendo, ramificándose y explorando el sustrato.
Las hifas tienen de 5 a 15 µm de diámetro y una longitud muy variable. Pueden estar tabicadas y, en ocasiones, los tabiques de separación están perforados. La zona apical es el extremo por el cual las hifas crecen y se extienden por el sustrato. Debido a esta actividad, la pared es más delgada y el citoplasma es denso con numerosas vesículas que contienen las sustancias necesarias para la formación de las nuevas estructuras que se requieren durante el crecimiento. En las hifas se distinguen:
· La pared que da forma a las hifas y les proporciona consistencia y resistencia a los cambios de presión osmótica. Está formada por dos tipos de componentes:
- Componentes fibrilares, principalmente quitina, un polímero de la N-acetilglucosamina.
- Componentes matriciales, sobre todo polisacáridos que rellenan huecos y dan grosor.
Hay hongos unicelulares que no presentan pared como consecuencia de su adaptación a la vida como parásitos endocelulares.
· La membrana plasmática está íntimamente ligada a la pared. En algunos casos forma invaginaciones con aspecto de vesícula.
· El citoplasma contiene los orgánulos característicos de las células eucariotas: mitocondrias, retículo endoplasmático, etc.
En el citoplasma también se presentan inclusiones diversas como gránulos de sustancias de reserva, principalmente de glucógeno y de esterol.
· Los núcleos son pequeños y, a menudo, haploides.
Los micelios de algunos hongos pueden modificarse para aumentar su poder de colonización, como en los cordones miceliares y en las estructuras rizomorfas.
Los cordones miceliares son agrupaciones de hifas que, en lugar de crecer ramificándose, crecen en paralelo.
En las estructuras rizomorfas, las hifas se desarrollan de modo que recuerdan a la raíz de una planta superior; en ellas se puede distinguir claramente el extremo por donde crecen.
|
|
|
|
|
Célula de levadura |
||

Con estas modificaciones los micelios consiguen avanzar de una manera más directa hacia un sustrato, al mismo tiempo que incrementan su capacidad de absorción de nutrientes.
Estructura levuriforme
Algunos grupos de hongos tienen, de manera permanente, una estructura unicelular, también denominada levuriforme. Habitualmente se les conoce como levaduras.
La célula de las levaduras tiene forma ovalada; hay especies con pared celular y especies que no la presentan.
Las células poseen el resto de los orgánulos propios de una célula eucariota. Como inclusiones suele contener materiales de reserva como lípidos y polifosfatos.
Las células pueden ser haploides o diploides, estas últimas, algo mayores y más activas que las células haploides.
En ocasiones las células aparecen unidas formando cadenas cortas.
Ciertas especies de hongos presentan estructura levuriforme sólo en alguna fase de su ciclo biológico.
1.2.2.2. Características funcionales: tipos de nutrición.
Son muy diversos los tipos de nutrición de estos grupos por eso los estudiaremos por separado.
BACTERIAS:
Las bacterias constituyen el único grupo de seres vivos que poseen representantes de todos los tipos de nutrición.
En el cuadro siguiente, recordamos los distintos tipos de nutrición y citamos un ejemplo de bacterias que los llevan a cabo.
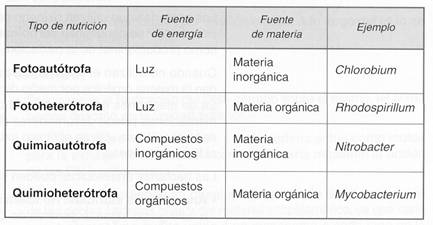
- Las bacterias fotoautátrofas utilizan la luz como fuente de energía, como el caso de las bacterias que poseen clorosomas con pigmentos capaces de realizar la fotosíntesis. La fuente de materia más frecuente es el CO2 que, al igual que las plantas, captan de la atmósfera e incorporan al ciclo de Calvin. También pueden utilizar el azufre y compuestos de azufre (S, H2S), o amoníaco, NH3, para obtener las moléculas necesarias para el metabolismo.
- Las bacterias fotoheterátrofas utilizan también la luz como fuente de energía, aunque la fuente de materia no es el CO2, sino compuestos orgánicos, en su mayoría glúcidos.
- Las bacterias quimioautátrofas oxidan diversos compuestos inorgánicos (NO2-, NH3, H2S). Como fuente de materia pueden tener diversos sustratos (CO2, S, H2S, NH3), al igual que las bacterias fotoautótrofas.
- Las bacterias quimioheterótrofas usan glúcidos, lípidos y proteínas para la obtención de energía y de materia. Algunas especies quimioheterótrofas colonizan a otros seres vivos, de los que obtienen los nutrientes. En ocasiones, esta relación beneficia tanto a las bacterias como al organismo al que colonizan, otras veces lo perjudican, como es el caso de las bacterias que provocan enfermedades.
También pueden ser descomponedoras de los restos de materia orgánica, lo que les convierte en microorganismos con un importante papel ecológico.
Las bacterias que utilizan oxígeno para degradar la materia y la obtención de ATPse denominan aeróbicas. Estas bacterias desprenden CO2 como producto final de la oxidación.
Cuando no utilizan el oxígeno, las bacterias son anaeróbicas. Degradan la materia orgánica por medio de fermentaciones, en las que el aceptor de electrones es una molécula orgánica. En algunos tipos de fermentaciones puede desprenderse dióxido de carbono. Algunas bacterias anaeróbicas utilizan nitritos o azufre como aceptores de electrones, al final del proceso.
Las bacterias anaeróbicas pueden clasificarse en:
- Anaeróbicas estrictas. No toleran el oxígeno y mueren. Pertenecen a este grupo algunos géneros de Clostridium.
- Anaeróbicas facultativas. Su metabolismo depende de la presencia de oxígeno; si hay oxígeno en el medio lo utilizan, si no, pueden prescindir de él y utilizar otro aceptor de electrones. Escherichia coli pertenece a este grupo.
- Aerotolerantes. No utilizan el oxígeno pero toleran su presencia. Streptococcus thermophilus es una bacteria de este tipo.
ALGAS:
Todas las algas tienen una nutrición autótrofa fotolitotrofa, aunque utilizan distintos pigmentos además de las clorofilas para captar la luz, según el grupo de algas.
PROTOZOOS:
Los protozoos son básicamente organismos heterótrofos. Distinguimos dos tipos de nutrición heterótrofa:
- Nutrición saprotrófica. Tiene lugar cuando los protozoos absorben sustancias disueltas en el medio. Estas sustancias pueden atravesar la membrana por difusión simple, por transporte activo o por pinocitosis, dependiendo del tamaño de las sustancias que se incorporan.
- Nutrición fagotróficau holozoica. Tiene lugar cuando los protozoos se alimentan de partículas o de otros microorganismos.
Si se alimentan de partículas pequeñas o bacterias, los protozoos se denominan micrófagos. Cuando se alimentan de partículas grandes, como fragmentos de vegetales u otros protozoos, se denominan macrófagos.
En este caso el alimento puede incorporarse a la célula por diversos mecanismos:

Con el alimento incorporado del exterior, los protozoos forman una vacuola digestiva.
A esta vacuola se vierten los enzimas digestivos, contenidos en los lisosomas, que degradan las sustancias alimenticias.
Cuando la digestión ha concluido, los nutrientes salen de la vacuola digestiva y pasan al citoplasma, en forma de pequeñas gotas. Por esta razón, la vacuola se va reduciendo de tamaño y en ella pueden quedar restos indigeribles que se expulsan por exocitosis, a través de cualquier punto de la membrana plasmática o bien por un orificio especializado denominado citopigio.
Los grupos que presentan nutrición autótrofa pueden realizar la fotosíntesis gracias a la presencia de cromoplastos, que contienen clorofilas y otros pigmentos amarillos y pardos.
De la fotosíntesis obtienen glucosa que transforman en almidón y en algunos casos otras sustancias de reserva como paramilo, leucosina o lípidos.
HONGOS:
Todos los hongos se nutren a partir de materia orgánica presente en el medio o en otros seres vivos.
Los hongos saprófitos aprovechan la materia orgánica procedente de restos de vegetales o de animales.
Los hongos parásitos utilizan la materia orgánica de organismos vivos.
Debido a esta capacidad para degradar sustancias procedentes de otros seres vivos, tienen gran importancia en el reciclaje de materia en los ecosistemas.
En los hongos verdaderos, la captación del alimento más característica tiene lugar por digestión externa: las hifas vierten al exterior enzimas que digieren el alimento y, posteriormente, absorben el producto de esta digestión. A continuación las hifas crecen y se extienden hacia una nueva fuente de alimento.
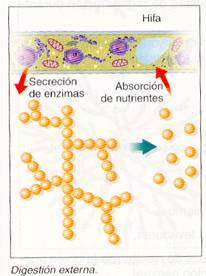
Las hifas crecen a una velocidad entre 1 y 4 mm/h. El crecimiento se produce por la combinación de las características de la pared en la zona apical, que es más delgada y plástica, y el empuje que produce la presión de turgencia sobre ella.
Los hongos parásitos desarrollan formas de crecimiento especiales que facilitan la infección.
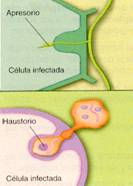
En los hongos parásitos, son muy comunes los apresorios. Éstos son inflamaciones de las hifas que presionan sobre las paredes de las células a las que infectan. Al mismo tiempo, segregan diversos tipos de enzimas que facilitan la penetración de las hifas en el interior de la célula.
En otros grupos se producen haustorios, que se introducen en el interior de las células sin perforar la membrana plasmática.
En ausencia de oxígeno, muchas especies de hongos verdaderos producen fermentaciones. Las fermentaciones tienen lugar en el interior de la célula; por ello utilizan sustratos que puedan ser absorbidos por éstas, como es el caso de moléculas de glúcidos de pequeño tamaño. El resultado de estas fermentaciones es la obtención de sustancias como acetaldehído o alcohol etílico.
Las levaduras tienen un metabolismo predominantemente fermentativo y se han utilizado desde hace mucho tiempo en la elaboración de productos alimenticios como el pan, el vino y la cerveza.